|
版权©1997年由美国生态学会
Keitt, T.H.、D.L.城市和B.T.米尔恩》1997。检测关键尺度分散的风景。保护生态(在线)1(1):4。从互联网上可用的。URL: http://www.consecol.org/vol1/iss1/art4/
本文在文本、数据表和附录是单独的文件可能被发现遵循这一点链接。
研究检测关键尺度分散的风景蒂莫西·h·Keitt1、2院长l .城市3布鲁斯·t·米尔恩1
1新墨西哥州大学生物学系;2圣菲研究所;3学校的环境中,莱文科学研究中心,杜克大学
关键词:连接,在分散的景观保护,分散,分散,栖息地连接与传播距离,景观,景观图、metapopulation,渗流,量化的栖息地连接在多个尺度,“垫脚石”补丁,思occidentalis最亮的星。
景观连通性不单独依赖规模;栖息地的配置或空间布置景观也是一个重要的行列式连接(福尔曼Baudry 1984年,加德纳et al . 1992年,Henein梅里厄姆1990年,泰勒et al . 1993年)。异质性的生境质量、贴片形状和大小的差异,和隔离的变化以及补丁导致景观空间变化连接(Gustafson和加德纳1996)。因此,尽管在整个景观的景观连通性指数平均对生态系统中比较有用,这些指标提供难以了解当地景观的连接结构。充分描述景观的空间结构,有必要考虑连接措施与当地相关地区和个人生境斑块。正如景观连通性的总体措施规模依赖,本地的连接可能会改变。
在这里,我们提出一个多尺度景观连通性的分析,基于统一的渗流理论的延伸(Stauffer Aharony 1985年,加德纳et al . 1987年,古尔德1988年Tobochnik, Creswick et al . 1992年)不均匀景观图(1993年坎特维尔和福尔曼,Harary 1969)。我们开发的总体措施景观连通性和patch-based措施个人贡献整体连通性。我们检查的敏感性景观连通性变化的景观配置,和敏感性和规模之间的关系。最后,我们考虑我们的分析的影响关于濒危物种栖息地的保护努力。
景观数据 我们的研究网站由一个1.5 x 106公里2美国西南部的地区,包括美国亚利桑那州科罗拉多、新墨西哥和犹他州。西南地区地形学的主要是由沙漠、岩石峡谷、半干旱草原、林地pinon-juniper,点缀着无数山脉支持针叶树和落叶阔叶林和麦克劳克林(1986)。从数字覆盖西南的地图(埃文斯等人。1993年,埃文斯和朱1993),我们选择两种森林覆盖,mixed-conifer杰克松,代表感兴趣的栖息地的模式;所有其他覆盖类型被认为是“nonhabitat。”The mixed-conifer habitat type was principally composed of Douglas-fir (Pseudotsuga menzesii),白冷杉(冷杉属concolor)和若干种松树(西黄松,p . contorta)。杰克松栖息地类型是由杰克松(p .杰克),但也可能包括一些pine-oak林地。1公里的栖息地地图有一个网格单元分辨率x 1公里。
我们最初动机选择杰克松和mixed-conifer森林类型的分布近似墨西哥斑点猫头鹰(适合筑巢栖息地思occidentalis最亮的星在美国西南部)。数据被用于开发一个复苏计划的墨西哥斑点猫头鹰(Keitt al.1995, USDI鱼类和野生动物服务1995)。
景观图 因为我们的研究关注景观斑块之间的空间关系,我们选择栖息地分布表示为一个数学“图”(Harary 1969)。图是由套点(顶点)和线(“边缘”)连接顶点。以前的工作表明图是一个有用的代表景观结构(坎特维尔和福尔曼1993),但依靠纯粹主观赋值顶点和边的景观功能。
我们开发了一个客观的手段产生景观图(图1)。输入数据由一个光栅栖息地地图每个网格单元分配一个价值如果细胞与选择的栖息地类型,如果细胞被认为是nonhabitat和零。一个补丁被定义为一组空间连续的栖息地的细胞。两个栖息地的细胞被认为是在同一生境补丁如果他们相邻的四个方位或对角线(东北、西北、等等)。一旦栖息地斑块标记,我们一个图的顶点分配给每个补丁。顶点之间的边被如果相应的补丁是根据标准来描述连接。因此,景观图包括一组顶点,每个栖息地的一个补丁,和一组边缘显示斑块之间的联系。用于显示目的,顶点被放置在每个补丁的几何中心。
图1所示。建设景观图。这个例子景观包含三个独立的栖息地生境斑块和一个集群,或“子图。“三个补丁属于单个集群,因为存在一个路径沿着图边缘(实线)连接所有三个补丁。如果任何一个显示的边缘被移除,将会有两个栖息地集群。如果所有边缘被移除,那么会有三个栖息地集群,每个组成的一个补丁。

我们最初的标签产生的森林数据> 6000个人生境斑块,小至一个网格单元。我们担心最小的补丁可能不代表真正的生境斑块,而是引入的噪声分类错误。因此,我们开发了一种新方法筛选潜在的分类错误的数据。我们的方法是基于应用中性模型(加德纳et al . 1987年)把补丁分成两组:那些有可能由于随机误差和那些更有可能代表真正的生境斑块。
在分类景观数据,小生境斑块更可能是随机误差的产品比大补丁。例如,如果错误是引入一个图像的概率p< < 1,那么创建一个块的错误的概率大小n大约是pn。随机误差是不可能产生大的补丁,因为n的增加,pn变得非常小。森林覆盖数据~地图面积的38%。的补丁中确定森林数据,大约50%是单细胞补丁。我们计算出一个随机地图覆盖18%也会有50%的单细胞补丁。因此,18%的错误率足以占所有单细胞补丁(即。,那些最有可能的结果分类错误)在森林里的数据。
我们接着问:块大小是什么频率的森林补丁类似补丁由随机噪声的期望频率?我们策划的频率森林补丁大小的块大小和观察到的随机地图密度为18%(100年图2),发现这两个分布是相似的一个补丁大小为10公里2。森林补丁> 10公里2大小发生在更高的频率比同尺寸的补丁在随机地图。因此,我们只保留补丁> 10公里2在随后的分析。
图2所示。块大小的相对频率西南部森林100年数据和随机地图密度为18%。补丁的补丁的大小增加频率衰减迅速随机地图,但森林数据的更慢。森林补丁> 10公里2大小发生的频率远高于随机地图,并因此可能导致分类错误。

扩展分析 为了探索规模效应景观连通性,我们定义一个补丁的连接标准基于补丁之间的最小安全距离。我们推断,传播的主要障碍是动物栖息地分布在这方面的差距会前往邻近的补丁。这些壁垒限制动物运动的程度,因此,连接,当然,取决于有机体感知景观格局的规模。影响物种能够长距离传播会减少差距与短程扩散比将物种栖息地分布。模仿分散行为的影响景观连通性,我们构造景观图通过将补丁之间的边缘只有在它们之间的最小距离小于指定阈值。的补丁集加入了边缘形成或孤立的组件图的子图。通过改变阈值的距离,我们能够解决的连接结构在各种尺度景观。
连接的措施 渗流理论是研究随机生成的连接结构(Stauffer和Aharony 1985)。因此,连接措施开发的渗流理论是一个自然的选择作为景观连通性的措施。大多数应用渗流理论生态学(加德纳et al.1987)是基于晶格渗流,细胞在晶格中占据的概率p和剩下的空置的概率1 -p。标准的数学导致晶格渗流是一大“跨越集群”的出现临界概率p暴击。如果相邻之间的连接,占据晶格细胞局限于只出现在北部,南部,东部,西部和方向,然后临界概率等于0.5928 (Stauffer和Aharony 1985)。其他重要概率可以通过改变连接规则,例如,通过允许连接到运行对角线(东北、西南等)之间的细胞。
渗流不需要经常发生晶格。一个更一般的模型包括渗流图结构,称为“债券渗透”(Stauffer和Aharony 1985)。因为正则格是可能的图形配置的子集(Harary 1969),债券自然渗透包括晶格渗流作为一个特例。债券渗流,网站要么是与概率有关p或与概率1 -无关p。在晶格渗流,大型集群连接的网站发生的临界值p。我们应用渗流理论代表了一个扩展的标准键渗流。在我们的模型中,两个生境斑块之间的连接的概率取决于补丁之间的距离。因此,跨越栖息地集群的出现发生在一个关键的意思是传播距离而不是在一个特定的栖息地的密度,如传统点阵渗流。渗流问题,连接概率不同景观被称为“梯度”或“非均匀渗透(米尔恩et al . 1996年)。
在渗流理论中,连接与连接集群的平均大小有关。自然的循环集群是其半径的大小。然而,在一般情况下,集群不圆;他们可以是不规则的,蜿蜒的结构。因此,衡量集群大小必须控制不规则形状。衡量集群大小用于渗流理论定义为“回转半径,”
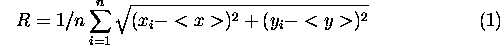
其中<x>和<y>的意思是x和y的坐标格子细胞集群中,x我和y我的坐标吗我th集群中的网格单元n是集群中的细胞的总数(Creswick et al . 1992年)。栖息地被定义为集群的补丁集阈值的子图或组件连接的景观图。几个生境斑块组成的集群,情商的总和。1接管所有栖息地细胞集群中的所有补丁。
与无单位的景观连通性指数,集群半径的单位是距离和直接的物理解释。想象一个集群随机移动粒子随机放在一个栖息地。回转半径是粒子的平均距离将之前遇到集群优势。类似地,如果一个分散动物仅限于移动集群(即在一个特定的栖息地。,它有一个低概率的遍历任何差距与另一个集群),其平均散布范围对应的半径集群。
size-weighted平均连通性的一组集群定义的相关长度的风景。给出一组集群的相关长度

在哪里米集群的数量和吗n年代是网格细胞集群的数量吗年代(Creswick et al . 1992年)。我们使用了相关长度作为一个总体衡量栖息地景观连通性。与回转半径、相关长度的单位是距离:个体的平均距离是可以分散在到达一个障碍,如果放置随机。因此,随着相关长度的增加,景观连通性增加。
补丁去除实验 我们不仅要衡量整体景观连通性,而且当地的补丁的连接措施。特别是,我们想知道影响景观连通性移除一个补丁的景观。我们设计了一个简单的分析,每个栖息地补丁是分别从景观中删除。我们记录的变化引起的相关长度消除每个补丁。因为删除补丁取决于规模的影响,除我们重复补丁分析大量的距离阈值。一片被移除后,建立了新的景观图。连通子图被识别和新的相关长度被记录。让C (d)景观图的相关长度,在距离阈值d,C (d,我)补丁后长度的关系我被删除,然后

规范化的重要性指数补丁吗我。规范化是一个相对重要性指数的贡献每个补丁整体景观连通性。我们选择了一个规范化的、无单位的指数,因为它是设计为among-patch比较精确。
我们预期的大补丁会有更大的重要性指标,只是由于它们的大小。因此,我们想要一个衡量单位面积的重要性。单位面积的重要性指数量化一个补丁的贡献单位面积上的整体景观连通性。我们单位面积指数定义为

在哪里n(我)栖息地的细胞的数量在补丁吗我和一个c网格单元的面积。1 /地区的单位面积指数单位,例如,公里2。
随机景观图 使渗流在景观图相当于债券渗流问题,有必要定义一个随机函数确定补丁之间的联系。因为成功的传播在栖息地分布差距意味着至少移动一段距离,如果不大于,补丁之间的最小距离,我们使用累积概率分布来表示两个补丁是否有效地连接对于一个给定的传播距离。阈值的风景图,散布函数仅仅是阶跃函数

在哪里d是补丁之间的最小距离。阈值函数的一个重要属性是一个给定的阈值距离导致只有一个景观图结构。然而,渗流问题可以推广到包括任何概率函数。超出了简单的阈值函数,概率累积负指数函数

在哪里p (d)是分散的概率至少距离d,k是分散系数。分散系数的单位是1 /距离;它的逆,1 /k分布下的平均传播距离。分布需要更多的参数,如伽马分布,可以使用。
不同阈值函数,负指数函数导致大量的景观图对于任何给定的扩散系数。标准技术在渗滤理论后,我们使用蒙特卡罗程序来生成多个景观配置。我们不同的扩散系数,k,从1.0(平均散布距离= 1公里)到0.01(平均散布距离= 100公里)。每个值的分散系数,100随机连接景观配置生成及其相关长度被记录。补丁在每种配置之间的连接由比较0.0之间的一个随机数。和1.0补丁之间传播发生的概率,根据情商。6。如果随机数不到传播的概率,补丁被加入到相同的子图或集群。在许多随机配置,两个补丁被加入的期望频率等于成功传播的概率。补丁远很少是连接,而斑块的边界常常联系,十分接近的频率取决于分散系数。
阈值的风景图 最大interpatch传播距离增加,森林覆盖地图越来越连接并最终合并成一个单一的、大跨越整个栖息地分布(图图3)。在从阈值距离的景观,在很大程度上是由独立的补丁和小生境集群。对于生物体能够分散20公里,景观是高度分散。在40公里,形成更大的子图,但风景还是划分为若干个栖息地集群。40公里以上,大部分的栖息地分布有关。尽管大多数的栖息地也加入了50公里,只有一个大的子图之间的边缘存在在东北和西南的栖息地分布。单一的顶点两端连接边缘被称为“接合点,”,因为删除任何一个将平分图(Harary 1969)。的阈值距离80公里,高度互联、图,一般来说,有许多替代途径从任何一个补丁到另一个。
图3所示。阈值的景观图。优势两个生境斑块之间绘制一条显示斑块之间的最小距离小于或等于给定的阈值距离。
点击每个图片查看详细。请注意详细的图像是非常大的。
10公里 20公里
20公里
30公里 40公里
40公里
50公里 60公里
60公里
70公里 80公里
80公里
90公里 100公里
100公里
突然从一个相对分离的栖息地分布主要连接地图作为一个拐点出现在相关长度的情节与阈值距离(图4)。40到45公里,景观的相关长度几乎翻了一番,从150公里增长到~ 275公里。突然连接在临界密度的变化,或在这种情况下一个阈值距离,在渗流理论被称为几何“相变”(Stauffer和Aharony 1985)。因此,生境斑块的景观是分为两个阶段:“相连接和断开连接的阶段。
图4所示。相关长度的栖息地分布与阈值距离。

景观敏感度 相关长度的变形不仅地图的连接和断开连接的阶段分离,但也与景观对生境变化的敏感度有关。补丁从地图中删除时,连接高度依赖规模效应(图5)。在比较短的距离阈值,删除一个补丁的影响很大程度上取决于它的大小,虽然形状也可能是重要的。阈值在0千米距离(所有补丁独立),图像敏感性主要是最大的补丁。
图5所示。在不同的阈值距离图的敏感性。补丁命令按大小。敏感性相关长度的变化,当一个补丁了。

在长阈值距离(30到40公里),最大的几个补丁引起的大偏差相关长度时删除。在30 - 40公里疏散距离,第二和第三大补丁由大量的图。到目前为止(0-40公里),图敏感性主要是最大的补丁。因此,补丁的相对位置和姿态很重要:补丁大小最重要的因素决定的重要性。
景观敏感度的模式改变了突然45岁和50公里。尽管最大的补丁仍然相对重要性指数高,敏感性(图图5)表现出几个大峰值产生的较小的补丁。大敏感性与这些补丁并不是由于他们的尺寸;相反,它反映了他们的角色为“垫脚石”补丁连接大面积的栖息地。这些垫脚石补丁与清晰度分图。
在距离过渡(60 - 80公里),以上图敏感性回到模式最大的补丁是最重要的。然而,整体灵敏度要低得多,特别是在80公里,因为阈值距离增加,替代途径之间的任何两个补丁的数量增加,并取消任何一个补丁对连通性影响很小。
补丁的重要性 个人的贡献补丁景观连通性测量通过删除每个补丁和记录相关长度的变化。因为景观的连通性是最敏感的补丁清除附近的尺度渗流转变,我们首先分析了景观的阈值距离45公里(图6)。正如预期的那样,最大的补丁已经连接的最大贡献。然而,当加权的区域,许多小补丁也大(单位面积)连接。这些补丁垫脚石补丁连接对应更大领域的栖息地。
图6所示。补丁的连接措施重要性45公里距离阈值。(左:我(45公里,我);正确的:一个(45公里,我)]。
点击每个图片查看详细。请注意详细的图像是非常大的。


因为经常不确定性的规模是最适合分析连接,平均连接在一系列尺度提供了一个更健壮的补丁的重要性。补丁重要性值平均阈值距离从0到100公里,在汉诺威的增量(图7),导致一个模式类似发现在45公里的阈值距离。然而,平均重要性指数包括更广泛的补丁,对应于45公里以外的地方渗流转换尺度。
图7所示。补丁的连接措施重要性平均阈值距离从0公里到100公里。(左:我(0 -100公里,我);正确的:一个(0 -100公里,我)]。
点击每个图片查看详细。请注意详细的图像是非常大的。


在随机连接景观图 尽管最大传播距离的假设是用于描述在不同的尺度景观的连通性,它不考虑传播事件的随机性质。因此,我们采用蒙特卡罗分析,基于概率分布函数,描述不同的连接性观察分散系数(见Eq。6)。
当扩散系数之间的不同k= 0.01(即。、远程传播)k= 1.0(即。短程扩散),传播行为之间的关系和景观的连通性是深刻的非线性,表现出强烈的变形在中间值(图8)。此外,相关长度的分布明显分为两个截然不同的阶段,高值指示连接阶段,较低的值不相交的阶段。在中间值,这两个阶段中存在相同的值的分散系数。因此,相变之前观察到最大的传播距离,在蒙特卡罗分析,直接关系到一个特定的分散系数。西南地区森林栖息地,临界分散系数(kc)是-0.07 ~ 0.06,即。14至17公里,平均传播距离。
图8所示。景观连通性为不同的扩散系数。系数是k在p (d)=ekd在哪里p (d)是单个分散的概率至少距离d。每个圆圈代表一个随机的景观配置。

连接性对于一个给定的范围扩散系数随着传播距离增加也表现出显著差异。两端的情节(k= 0.01,k= 1.0),连接的范围值是最小的。远程传播导致了景观连接每蒙特卡罗迭代。短程扩散,风景总是支离破碎。在中间传播系数,附近的渗流转变,连接值的范围更大。大范围的连接值再次反映了相变附近发现敏感性增加。
我们的分析的一个重要发现是,栖息地的丧失一个高度依赖规模影响景观连通性。感知的生物景观在细尺度、景观配置和垫脚石补丁的后果很小,因为当地人口限制在生境斑块。同样,运动的物种能够远程传播不会强烈影响的配置单独的补丁。对于物种附近的渗流转变、景观配置可能扮演了一个重要的角色在决定景观连通性。附近的渗流转变,个别补丁可以作为走廊或垫脚石,弥合差距的栖息地分布。因此,我们认为,景观配置将重要物种的传播行为的地方风景渗流阈值附近。
这些结果适用于不同物种如何在某种程度上取决于所使用的传播方式。我们设计分析捕捉基本特征“sky-island栖息地之间的猫头鹰分散西南的特征。使用其他物种的传播模式将与不同的庄园景观模式。有机体必须步行或运行在一个景观可能遇到的障碍不代表我们的栖息地地图。然而,我们没有理由不能修改我们的方法将其他传播方式和更详细的空间信息。
一般来说,我们有太少的数据可以精确预测墨西哥斑点猫头鹰的传播模式。然而,有几个报告(综述Keitt et al . 1995年)青少年猫头鹰分散> 45公里,是典型的,至少在一些地区猫头鹰的范围,为青少年建立新界10 - 20公里从他们出生的领土(Keitt et al . 1995年)。因此,我们认为,墨西哥斑点猫头鹰可能存在metapopulation,景观连通性是一个重要的考虑他们的生存。
管理的角度 应用程序对地区级的核生境分析保护生态代表向前迈出了重要一步,因为它的地方在景观环境管理决策,从而避免当地政策,未能识别关键种群之间的联系。特别是,我们相信补丁重要性指数(无花果。和7)可能有助于指导栖息地保护的努力和指定关键栖息地,以及传播和人口研究的设计。例如,墨西哥斑点猫头鹰的在我们的研究中,一个栖息地面积、泰勒山管理员在新墨西哥州中部地区,有很高的重要性在所有分析指标。泰勒山区域以前是不被认为是重要的栖息地,因为它包含相对较少的猫头鹰。然而,我们的分析强调了泰勒山作为跳板链接猫头鹰种群在新墨西哥州和亚利桑那州南部人口在科罗拉多州和犹他州。这些北方人群可能存在人口下沉(Pulliam 1988),但是可以作为缓冲对源种群崩溃造成的干扰或疾病(托马斯·et al . 1996年)。
区域和景观尺度分析的重要性更大的情况下,多个机构管理在一个物种的栖息地范围。例如,市场力量可能引起森林经理收获木材从一个小地区的栖息地,如泰勒山认为小生态重要性的补丁。然而,高单位面积指数(图6)同样的补丁可能导致经理承认补丁的作用是一个重要的链接在一个更广泛的地区由几个管理辖区。补丁基于非空间,经济视角的改变可能危及众多机构保护目标。管辖范围内(如国家森林,经理可能会重新分配收成补丁,有小单位面积指数为了保持连接整个栖息地的价值网络。或者,一个或多个机构可能贸易土地或提取的网络资源来实现管理目标,同时保持忠诚。
我们还想强调,我们的研究结果并不局限于单一物种的研究。生境斑块,有很高的重要性在一个广泛的尺度(图7)可能代表重要传播许多物种的栖息地。multipecies生态层面,分析的结果也可以由覆盖distance-percolation研究许多栖息地生境类型和组合类型。由此产生的地图栖息地的重要性可以被使用,以及适当的支持数据和研究,作为生态系统的基础管理。
传播、生物地理学和气候变化beplay竞技 渗流研究中一个有趣的假设提出我们的西南地区是否存在一种机制,行为或进化,这将导致传播距离之间的临界距离就在断开连接景观和景观。传播距离与尺度相关的临界长度可能会是一个最佳的传播成本之间的权衡,找到一个合适的可能性,无人占领的领土。因此,在某种意义上,人口分散的自然栖息地可能会发展向metapopulation动力学。
重要的传播也暗示了原点和维护生物多样性。例如,在莱特(1970)的平衡变化理论,断断续续的孤立的种群之间的基因流是一个重要的机制允许人口个人探索许多遗传的可能性。这样一个机制,结合关键连接数量,可以解释哺乳动物的高多样性与山地环境在美国西部(克尔和封隔器1997)。
beplay竞技气候变化也可能发挥作用,在多样性的起源和隔离和濒临灭绝的物种。通常,预计树分布在海拔上升,气候变暖,从而消除峰会栖息地的面积和降低高海拔中期植被类型。转变,减少栖息地面积可能产生物种灭绝Lomolino et al . (1989)。此外,高程变化应该改变网络几何和补丁需要分散为更远距离的有效链接。额外的本地人口灭绝或碎片可能发生如果合成临界距离超过一个特定物种的平均分散能力。因此,气候变beplay竞技化和土地利用实践,改变补丁网络几何图形可能与传播距离,从而阻碍了一些物种而促进他人。我们看到一个开放的和有趣的研究领域研究中景观图是如何随时间而变化的气候和地形的变化,以及这些变化与物种灭绝和物种多样性。
本文对被邀请。如果发表,你的反应将是超链接的文章。提交评论,跟进这个链接。阅读评论已经接受,跟进这个链接。
确认
我们希望感谢a·约翰逊建议和评论这个手稿的早期草稿。我们感谢墨西哥斑点猫头鹰康复团队的成员——B。块,g .白色,a .富兰克林·沃德s Rinkevitch闪烁发光,和j . Ganey——三个匿名评论者批判性审查这项工作。我们感激承认美国鱼类和野生动物服务的支持物和在野势力的合作协议。1448-00002-94-0810)。
- Cantwell教授,m D。,R.T.T. Forman.1993年。景观图:生态建模与图论来检测配置多样化的景观。景观生态学 8(4):239 - 251。
- Creswick, r . J。、H.A. Farach C.P.普尔,Jr。1992年。介绍重正化群方法在物理学。约翰威利,纽约,纽约,美国。
- 埃文斯,d . L。,Z. Zhu.1993年。AVHRR森林映射:国家应用和全球影响。76 - 79页在a·刘易斯,编辑器。展望未来着眼于过去。学报1993 ACSM / ASPRS公约美国,马里兰州贝塞斯达ASPRS,。
- 埃文斯,d . L。,朱z, k . Winterberger。1993年。森林分布与AVHRR数据的映射。世界资源评估 5:66 - 71。
- 福尔曼,r·T·T。和j . Baudry。1984年。在景观生态学树篱和灌木篱墙网络。环境管理 8:495 - 510。
- 加德纳,r . H。B.T.米尔恩,随机变数奥尼尔,M.G.特纳。1987年。中性模型的分析大规模景观模式。景观生态学 1:19-28。
- 加德纳,r . H。随机变数,奥尼尔、M.G.特纳和V.H.戴尔。1989年。量化尺度效应与简单的渗流模型动物的运动。景观生态学 3:217 - 227。
- 加德纳,r . H。M.G.特纳,随机变数奥尼尔,美国Lavorel。1992年。模拟的尺度景观边界对物种的影响持久性和传播。76 - 89页在m . m .荷兰、p·g·里瑟尔和r . j .乃编辑。景观边界的角色在改变环境的管理和恢复。查普曼和大厅,纽约,纽约,美国。
- 古尔德,H。,J. Tobochnik.1988年。介绍计算机仿真方法:应用物理系统。addison - wesley、阅读、麻萨诸塞州,美国。
- Gustafson, e . J。,R.H. Gardner.1996年。景观异质性的影响在补丁殖民的概率。生态 77年(1):94 - 107。
- Harary F。1969年。图论。addison - wesley级数在数学。addison - wesley、阅读、麻萨诸塞州,美国。
- Henein, K。,G. Merriam.1990年。的元素连接在走廊质量是可变的。景观生态学 4:157 - 170。
- 约翰逊,a。R。B.T.米尔恩,J.A.恩斯。1992年b。在分形扩散景观:tenebrionid甲虫运动的模拟和实验研究。生态 73年(6):1968 - 1983。
- 约翰逊,a。R。克里斯特恩斯j, B.T.米尔恩,世贸组织。1992年一个。动物运动和种群动态异构风景。景观生态学 7(1):63 - 75。
- Keitt, t·H。a·富兰克林,D.L.城市。1995年。景观分析和metapopulation结构。第三章,墨西哥斑点猫头鹰的恢复计划,卷二:技术和支持信息。美国内政部鱼类和野生动物服务,西南地区,美国新墨西哥州阿尔伯克基。
- 克尔,j . T。,L. Packer.1997年。生境异质性的行列式在高能地区哺乳动物物种丰富度。自然 385年:252 - 254。
- Lomolino, m V。,J.H.棕色和r·戴维斯。1989年。岛屿生物地理学的山地森林哺乳动物在美国西南部。生态 70年(1):180 - 194。
- 麦克劳克林,s P。1986年。美国西南部的植物区系分析。大盆地博物学家 46(1):46 - 65。
- 梅里厄姆,G。1984年。连接:一个基本的生态景观格局的特点。页面5-16在j·布兰德和p·阿格,编辑。第一届国际研讨会论文集在景观生态研究和规划方法。我:主题景观生态的概念,卷1。鲁开德大学中心图书公司,鲁开德那样,丹麦。
- 米尔恩,b . T。境约翰逊,T.H. Keitt c.a哈特菲尔德,大卫j . p Hraber。1996年。检测与pinon-juniper林地群落交错区相关的临界密度。生态 77年(3):805 - 821。
- 诺,r F。1991年。景观连通性:不同的功能和不同的尺度。页面27-39在w·e·哈德逊,编辑器。景观和生物多样性的联系。岛出版社,华盛顿特区,美国。
- ·普廉姆h·R。1988年。源、汇、和人口的监管。美国博物学家 132年(5):652 - 661。
- 香农,c, E。,W. Weaver.1949年。通信的数学理论。伊利诺伊大学出版社,厄巴纳伊利诺斯州,美国。
- Stauffer D。,一个。一个harony.1985年。介绍了渗流理论。泰勒和弗朗西斯,伦敦,英国。
- 泰勒,p D。,Fahrig . k . Henein和g·梅里厄姆。1993年。景观结构的连接是一个至关重要的元素。Oikos 68年(3):571 - 573。
- 托马斯,c, D。贝拉的歌手,D.A.理查德。1996年。蝴蝶metapopulation灾难性灭绝人口来源。美国博物学家 148年(6):957 - 975。
- 美国内政部,鱼类和野生动物服务。1995年。墨西哥斑点猫头鹰的恢复计划:美国新墨西哥州阿尔伯克基卷。。
- 赖特,S。1970年。进化的随机漂移和转移平衡理论。在k .小岛编辑器。在群体遗传学数学主题。斯普林格出版社,德国海德堡。
记者的地址:
蒂莫西·h·Keitt
圣菲研究所
海德公园路1399号
美国新墨西哥州圣达菲87501
电话:505-984-8800
传真:505-982-0565
tkeitt@santafe.edu
本文对被邀请。如果发表,你的反应将是超链接的文章。提交评论,遵循说明作者。
![]()