|
版权所有©1997韧性联盟*
路德维希,D. B.沃克和C. S.霍林,1997。可持续性、稳定性和弹性。保护生态(在线)1(1): 7。可从互联网上获得。URL: http://www.consecol.org/vol1/iss1/art7/
本文的一个版本,其中的文本、图、表和附录是单独的文件,可以通过以下命令找到链接.
洞察力可持续性、稳定性和弹性唐纳德·路德维希1,布莱恩•沃克2,克劳福德s温和3.
1英属哥伦比亚大学数学系;2CSIRO野生动物与生态学处;3.佛罗里达大学动物学系
关键词:分岔;多个稳定状态;弹性;稳定。
平衡 如果作用在机械系统上的力是平衡的,那么机械系统就处于平衡状态。例如,当一个物体漂浮时,重力被液体的位移产生的浮力所平衡。“自然的平衡”(皮姆1991)是这一思想在自然世界中的延伸。这个概念通常指的是能量和物质的稳定流动,而不是指一个系统的组成部分不发生变化。
弹性和稳定性 我们感兴趣的是描述具有弹性的自然系统,即在受到干扰时倾向于保持其完整性的自然系统(Holling 1973)。这与稳定性的概念有关。稳定的非正式概念是指系统在受到干扰时返回平衡状态的趋势。如果突然给漂浮在水面上的筏子加一个重物,通常的反应是使负重的筏子振荡,但振荡的振幅逐渐减小,因为振荡的能量以波的形式消散,最终以热的形式消散。加了重量的木筏与没有加重量的木筏会在不同的位置静止,但我们认为新的结构与旧的基本相同。系统稳定。
如果我们逐渐增加筏子的重量,最终结构会改变。如果重物挂在筏子下面,筏子就会陷得越来越深,因为需要越来越大的位移来平衡更高的重力。最终,浮力无法平衡重力,整个结构下沉:系统不再稳定。另一方面,如果重物放在木筏上,木筏可能会突然翻转,在整个系统下沉之前,重物和其他内容物就会消失。这种突然的失去稳定性可能比逐渐下沉更危险,因为几乎没有预警或准备的机会。我们可以认为筏子系统的弹性随着重量的增加而减弱。
假设我们接受“自然平衡”和它所暗示的资源的稳定流动。随着我们对自然系统产品的需求越来越大,我们又给它们装上越来越多的废物,我们可能会经历逐渐失去稳定性还是突然失去稳定性呢?为了澄清这些问题,我们必须改进我们的术语。为了决定一个系统是否稳定,我们必须首先指定配置中的更改或完整性的丧失是什么意思。如果我们不关心筏子在称重时是否翻转,那么浮筏就不存在突然失去稳定性的问题。我们还必须明确可能影响系统的干扰的类型和数量。假设一个固定的重物放在一个有人的木筏上。如果木筏上的人移动,木筏可能会以稍微不同的角度漂浮,但如果他们移动太远或同时移动,木筏可能会倾斜。不导致倾倒的乘客可能的运动范围被称为竖直状态的稳定域或吸引域。如果固定重量的量逐渐增加,平衡就变得更加不稳定,因此,吸引力的范围就会缩小。 Eventually, the weight becomes large enough so that there is no domain of attraction at all, and the raft will flip over no matter what its occupants do.
前面的例子区分了筏子的重量和乘员的位置。如果重量的变化非常缓慢或根本没有变化,我们可以认为“系统”是由木筏和重量组成的。居住者的位置变化相对较快,这些变化可以被认为是系统的干扰。另一方面,我们可以采取更全面的观点,把木筏、重量和乘员视为一个单一的系统。如果居住者组织自己来预测和纠正外部干扰,那么系统可能能够保持其完整性足够长的时间,让他们实现他们的目标。另一种可能的应对干扰的方法是重组木筏本身。如果它是由几个松散耦合的子单元构成的,那么过度的权重或强烈的扰动可能会翻转系统的一部分,但保留其余的完好无损。这样的结构可能不需要像单个木筏那么多的警觉性来维护,而且它可能能够承受更多的外部干扰。另一方面,如果连接子单元的绑定变得僵硬,那么结构可能变得脆弱,因此更容易失败。这个简单的例子说明了系统弹性的概念如何取决于我们的目标、感兴趣的时间尺度、扰动的性质和量级、系统的底层结构以及可行的控制措施的种类。
第二节提出了简单的一维原型系统的稳定性和弹性的主要思想。可以对这些原型系统进行显式计算,但它们的定性行为适用于更复杂的例子。第三节用云杉芽虫模型说明了双稳态平衡、硬失稳、滞回和弹性的概念。虽然这里没有尝试为这些系统提供一个正式的模型,但在budworm模型的行为和各种生态系统之间有质的相似性,特别是湖泊生态系统、波罗的海和北方森林。这样的模型是已知的第四节为了一个竞争性的放牧系统。该系统具有类似于一维模型的定性行为,它也表现出滞后和硬失稳。提出了一个类似的系统,涉及火作为调节过程第五节.后一种体系也表现出有规律的振荡。的附录给出了扰动系统的返回时间与其弹性之间关系的详细说明。弹性有两种相互冲突的定义,这可能会造成混淆。皮姆(1991)的定义只适用于线性系统的行为,或非线性系统在稳定平衡附近的行为,其中线性近似是有效的。在皮姆看来,弹性的丧失是由于接近稳定平衡的缓慢动力所致。我们在这里使用的Holling(1973)的定义是指一个非线性系统在吸引域边界附近的行为。在我们的意义上,弹性的丧失与划分吸引力区域的缓慢动态有关。
全球的稳定 自然平衡的概念可以理解为,在任何形式的扰动下,系统都将保持其完整性。当我们对自然系统进行重大修改时,可能会(通常是无意识地)做出这样的假设。我们的期望是,事情将或多或少地像以前那样进行,系统的响应将近似地与摄动成正比。这种行为可以用最简单的线性模型来表示。有些人可能会争辩说,节俭原则规定,在没有强有力的相反证据的情况下,应该使用这样的模型。下面的线性模型说明了全局稳定性的性质,这意味着系统将总是返回到某个平衡,不管它被移出该平衡有多远。
假设动力学是由形式的关系给出的
在哪里 是一个外部变量的平滑变化函数吗
是一个外部变量的平滑变化函数吗 而且
而且 是利息的数量。然后
是利息的数量。然后 如果
如果 ;系统在这里有一个平衡。这个平衡是稳定的,因为
;系统在这里有一个平衡。这个平衡是稳定的,因为 如果
如果 ,
, 如果
如果 .这些关系意味着系统接近平衡,无论起点是什么。
.这些关系意味着系统接近平衡,无论起点是什么。
一个系统,例如(情商。)不会让我们失望或惊讶。无论偏移多远,它都会回到平衡状态,并且平衡的位置会随着外生变量的变化而平稳变化 .这样的系统不适合讨论自然系统可能的崩溃,因为这种崩溃被下列假设排除在外(情商。).数学理论提供了许多不同行为的例子,我们的目标是研究它们的合理性。不幸的是,许多理论(包括大多数经济理论)都是基于类似于(情商。).特别是,Pimm(1991)从系统的角度定义了“弹性”(情商。),这种特别的假设可能会误导粗心的人。有一种数学理论表明,诸如(1)这样的系统是具有稳定平衡的一般系统的良好近似值,但该理论暗示,这种近似值只在平衡的直接附近成立,也就是说,这种近似值只在局部有效。详情载于附录。
.这样的系统不适合讨论自然系统可能的崩溃,因为这种崩溃被下列假设排除在外(情商。).数学理论提供了许多不同行为的例子,我们的目标是研究它们的合理性。不幸的是,许多理论(包括大多数经济理论)都是基于类似于(情商。).特别是,Pimm(1991)从系统的角度定义了“弹性”(情商。),这种特别的假设可能会误导粗心的人。有一种数学理论表明,诸如(1)这样的系统是具有稳定平衡的一般系统的良好近似值,但该理论暗示,这种近似值只在平衡的直接附近成立,也就是说,这种近似值只在局部有效。详情载于附录。
分岔 为了探究局部稳定性和全局稳定性之间的差异,我们必须研究非线性模型,即状态变量出现在比线性函数更复杂的函数中的模型。下面的例子有三个均衡,而不是一个。的三次或更复杂的依赖
变量,例如,
在这里,
 是一个参数或一个缓慢变化的量,其动态与当前无关。系统的均衡是
是一个参数或一个缓慢变化的量,其动态与当前无关。系统的均衡是 .这是两个因素中的任何一个(情商。)就消失了。因此,它们是
.这是两个因素中的任何一个(情商。)就消失了。因此,它们是
如果
 ,那么就有三个均衡
,那么就有三个均衡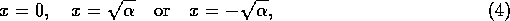
但是,如果
 ,则只有一个平衡点
,则只有一个平衡点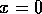 .均衡结构或稳定性的这种变化被称为“分岔”(Guckenheimer and Holmes 1983)。它意味着系统的定性行为的变化。为了探究这个特性,我们必须讨论一些附加的概念。
.均衡结构或稳定性的这种变化被称为“分岔”(Guckenheimer and Holmes 1983)。它意味着系统的定性行为的变化。为了探究这个特性,我们必须讨论一些附加的概念。
局部稳定和吸引域 为了确定平衡的稳定性,检查速度的符号就足够了
.例如,如果 ,第二个因素(情商。)总是积极的,因此,如果
,第二个因素(情商。)总是积极的,因此,如果 而且如果
而且如果 .在这种情况下,系统总是远离状态.我们得出的结论是平衡在是不稳定的,如果
.在这种情况下,系统总是远离状态.我们得出的结论是平衡在是不稳定的,如果 .另一方面,如果
.另一方面,如果 ,然后
,然后 改变三个地方的标志:
改变三个地方的标志:
平衡在哪里
 是不稳定的,因为系统总是远离那个点如果附近(根据方程式。而且6).同样的,平衡态
是不稳定的,因为系统总是远离那个点如果附近(根据方程式。而且6).同样的,平衡态 是不稳定的(根据方程式。而且8).另一方面,平衡是是稳定的(根据方程式。7),因为从附近点的运动是朝向那个点的。但是,如果系统在间隔时间之外启动
是不稳定的(根据方程式。而且8).另一方面,平衡是是稳定的(根据方程式。7),因为从附近点的运动是朝向那个点的。但是,如果系统在间隔时间之外启动 时,它会偏离平衡.因此,平衡在是局部稳定的,但不是全局稳定的。系统返回到如果是小扰动,但较大的扰动会使系统进入不稳定区域。的时间间隔
时,它会偏离平衡.因此,平衡在是局部稳定的,但不是全局稳定的。系统返回到如果是小扰动,但较大的扰动会使系统进入不稳定区域。的时间间隔 叫做“吸引域”的点是什么,因为在这个区间内开始的轨迹最终会回到而不是那些从户外开始的。
叫做“吸引域”的点是什么,因为在这个区间内开始的轨迹最终会回到而不是那些从户外开始的。
很明显,稳定平衡的吸引域在收缩, 减少对零。三个均衡崩溃为一个
减少对零。三个均衡崩溃为一个 时,只有一个不稳定平衡存在
时,只有一个不稳定平衡存在 .这些信息总结在图1.这个图看起来像一个分支,因此,它被称为“分岔图”。点的吸引域是包含在两个弯曲的分支中,没有其他的吸引域。
.这些信息总结在图1.这个图看起来像一个分支,因此,它被称为“分岔图”。点的吸引域是包含在两个弯曲的分支中,没有其他的吸引域。
图1.的参数
 在横轴上标出了对应的平衡
在横轴上标出了对应的平衡 为(2)式。都画在纵轴上。稳定平衡用实线表示,不稳定平衡用虚线表示。
为(2)式。都画在纵轴上。稳定平衡用实线表示,不稳定平衡用虚线表示。
关于自然界中存在多稳定状态的证据越来越多:珊瑚礁(Done 1992, Hughes 1994, McClanahan等人1996)、非洲牧场(Dublin等人1990)、浅湖(Schindler 1990, Carpenter和Leavitt 1991, Scheffer等人1993)、海带林(Estes和Duggins 1995)和草原(D’antonio和Vitousek 1992, Zimov等人1995)。
干扰和缓慢的参数变化
我们已经看到了 时,系统趋于稳定平衡如果是在吸引的范围内开始。如果我们设想干扰将系统移动一段距离
时,系统趋于稳定平衡如果是在吸引的范围内开始。如果我们设想干扰将系统移动一段距离 从稳定平衡,它们不会影响系统的完整性(其倾向于返回0状态),只要
从稳定平衡,它们不会影响系统的完整性(其倾向于返回0状态),只要 .现在,如果我们允许参数
.现在,如果我们允许参数 慢慢减少:慢慢地减少
慢慢减少:慢慢地减少 ,系统将需要越来越长的时间返回到状态当是流离失所,因为附近的运动非常缓慢
,系统将需要越来越长的时间返回到状态当是流离失所,因为附近的运动非常缓慢 ,以及震级的扰动可使系统进入慢动力学区域。我们可以考虑减少
,以及震级的扰动可使系统进入慢动力学区域。我们可以考虑减少 因为系统的完整性受到一定程度干扰的威胁越来越大。恢复力丧失的一个症状可能是恢复的时间越来越长在干扰。回归时间和弹性之间的联系并不完全直接;我们在附录中详细说明了这一点。
因为系统的完整性受到一定程度干扰的威胁越来越大。恢复力丧失的一个症状可能是恢复的时间越来越长在干扰。回归时间和弹性之间的联系并不完全直接;我们在附录中详细说明了这一点。
吸引力的两个领域 前面的系统对于自然系统来说不是一个可信的模型,因为它预测了状态变量在某些情况下可能趋于无穷。一种更合理的情况是系统可能从一个稳定均衡变为两个稳定均衡。接下来我们考虑一些这样的“双稳态”系统。
通过改变的符号,我们得到了这类系统的一个简单原型在(情商。).如果时间方向相反,则稳定平衡和不稳定平衡互换。如果 ,则单平衡在全局稳定:无论从哪里开始,系统总是返回到平衡状态。而不是两个不稳定均衡
,则单平衡在全局稳定:无论从哪里开始,系统总是返回到平衡状态。而不是两个不稳定均衡 (和前一种情况一样)会有两个稳定均衡。新制度是
(和前一种情况一样)会有两个稳定均衡。新制度是
如果 ,对于这个系统,我们有
,对于这个系统,我们有
如果最初,然后朝向平衡点 ,但如果最初,然后朝向平衡点
,但如果最初,然后朝向平衡点 .因此,点的吸引域
.因此,点的吸引域 是积极的轴,和的吸引域
是积极的轴,和的吸引域 是消极的轴。每个稳定均衡都是局部稳定的,但不是全局稳定的。这个系统可以从一个稳定状态翻转到另一个稳定状态通过越过不稳定线.因为这条线分隔了两个吸引域,所以被称为“分隔线”。的平衡在策划图2.
是消极的轴。每个稳定均衡都是局部稳定的,但不是全局稳定的。这个系统可以从一个稳定状态翻转到另一个稳定状态通过越过不稳定线.因为这条线分隔了两个吸引域,所以被称为“分隔线”。的平衡在策划图2.
图2.平衡在
为(9)式。绘制vs。 如图1所示。时间增加的方向与图1相反;因此,稳定平衡和不稳定平衡互换了。
如图1所示。时间增加的方向与图1相反;因此,稳定平衡和不稳定平衡互换了。
干扰和缓慢的参数变化
分岔图图2包含了大量关于系统对扰动的响应。如果 时,系统将回到稳定平衡无论干扰有多大,它都无处可去。然而,如果
时,系统将回到稳定平衡无论干扰有多大,它都无处可去。然而,如果 系统从较低的分支开始,如果移位少量,它会倾向于返回到那里。作为
系统从较低的分支开始,如果移位少量,它会倾向于返回到那里。作为 趋于零时,稳定平衡态和不稳定平衡态之间的距离减小。因此,给定量级的扰动使系统越来越接近不稳定平衡。在接近不稳定平衡时,动力学是缓慢的,因此,对于接近不稳定平衡的轨迹,返回较低分支附近的时间急剧增加。关于回程时间的这一点可以通过类似于附录中的计算来精确计算。
趋于零时,稳定平衡态和不稳定平衡态之间的距离减小。因此,给定量级的扰动使系统越来越接近不稳定平衡。在接近不稳定平衡时,动力学是缓慢的,因此,对于接近不稳定平衡的轨迹,返回较低分支附近的时间急剧增加。关于回程时间的这一点可以通过类似于附录中的计算来精确计算。
为了更大程度的干扰 时,可将系统移过分隔线,并可能接近稳定均衡的上支。在随机扰动模式下,我们可能会看到系统花很长时间在一个或另一个稳定平衡附近。不时地,这些随机扰动可能结合起来,使系统达到另一个稳定平衡。生态学家研究的“Allee效应”提供了双稳态系统的一个例子。一个种群在数量低的情况下可能会减少生存或繁殖成功率。例如,如果鱼群的数量相对于捕食者的能力足够高,那么鱼群的人均死亡率往往较低。如果这些鱼因捕鱼压力或环境恶化而数量减少,它们的数量可能会减少,最终在当地灭绝。另一方面,一个庞大的人口可以在很长一段时间内维持自己。这种动态可能解释了沙丁鱼和凤尾鱼之间偶尔的优势转换,正如在加利福尼亚海岸沉积的它们的鳞片所揭示的那样。类似的模式出现在地球物理特征中,如地球磁场的极性、涉及墨西哥湾流的海洋环流和气候波动,但其中一些波动可能太有规律,不完全是随机的。 For larger values of
时,可将系统移过分隔线,并可能接近稳定均衡的上支。在随机扰动模式下,我们可能会看到系统花很长时间在一个或另一个稳定平衡附近。不时地,这些随机扰动可能结合起来,使系统达到另一个稳定平衡。生态学家研究的“Allee效应”提供了双稳态系统的一个例子。一个种群在数量低的情况下可能会减少生存或繁殖成功率。例如,如果鱼群的数量相对于捕食者的能力足够高,那么鱼群的人均死亡率往往较低。如果这些鱼因捕鱼压力或环境恶化而数量减少,它们的数量可能会减少,最终在当地灭绝。另一方面,一个庞大的人口可以在很长一段时间内维持自己。这种动态可能解释了沙丁鱼和凤尾鱼之间偶尔的优势转换,正如在加利福尼亚海岸沉积的它们的鳞片所揭示的那样。类似的模式出现在地球物理特征中,如地球磁场的极性、涉及墨西哥湾流的海洋环流和气候波动,但其中一些波动可能太有规律,不完全是随机的。 For larger values of 在美国,人们会认为从一种平衡跳到另一种平衡是极其罕见的。因此,我们可以把……的增加联系起来
在美国,人们会认为从一种平衡跳到另一种平衡是极其罕见的。因此,我们可以把……的增加联系起来 随着韧性的增强。
随着韧性的增强。
Ludwig, Jones和Holling(1978)使用以下模型来理解云杉芽虫的动态。的数量 表示芽虫密度,以每英亩幼虫计算。这个密度假设随时间变化,根据
表示芽虫密度,以每英亩幼虫计算。这个密度假设随时间变化,根据
在哪里 是低密度下的内在增长率,
是低密度下的内在增长率, 是在没有捕食的情况下,budworm的一种承载能力,在(Eq。14)为捕食率。假设捕食者具有霍林iii型功能反应,其最大捕食率为
是在没有捕食的情况下,budworm的一种承载能力,在(Eq。14)为捕食率。假设捕食者具有霍林iii型功能反应,其最大捕食率为 和半饱和的budworm密度
和半饱和的budworm密度 .这一函数形式表明,在虫密度的中间范围内,捕食者对动态的影响最大。在密度较低的情况下,捕食者会寻找其他猎物,因为捕食budworm的回报相对较低。在高密度时,百虫会淹没它们的捕食者;因此,掠食者的人均效应很小,就像掠食者对大鱼群的人均效应很小一样。的参数
.这一函数形式表明,在虫密度的中间范围内,捕食者对动态的影响最大。在密度较低的情况下,捕食者会寻找其他猎物,因为捕食budworm的回报相对较低。在高密度时,百虫会淹没它们的捕食者;因此,掠食者的人均效应很小,就像掠食者对大鱼群的人均效应很小一样。的参数 是与树叶密度成正比的,因为捕食者在树叶中寻找百虫,它们的反应是由每单位树叶中百虫的数量调节的。因此,
是与树叶密度成正比的,因为捕食者在树叶中寻找百虫,它们的反应是由每单位树叶中百虫的数量调节的。因此, 实际上是一个状态变量它的变化时间比虫的慢。目前,我们关注
实际上是一个状态变量它的变化时间比虫的慢。目前,我们关注 是一个常数。
是一个常数。
Ludwig et al.(1978)提供的一些代数表明,根据无量纲参数的大小,有两个或四个budworm的平衡 而且
而且 ,由
,由

这些平衡满足
在哪里 .平衡
.平衡 总是不稳定的,因为
总是不稳定的,因为 如果
如果 是小而正的。最高平衡总是稳定的,因为
是小而正的。最高平衡总是稳定的,因为 如果是非常大且正的。因此,如果只有两个平衡态,虫密度总是向上层平衡态移动。当有四个平衡时,它们的稳定性交替变化。一个典型案例见图3.如果之间的是
如果是非常大且正的。因此,如果只有两个平衡态,虫密度总是向上层平衡态移动。当有四个平衡时,它们的稳定性交替变化。一个典型案例见图3.如果之间的是 而且
而且 ,可能接近高平衡或低平衡,取决于起始位置是否在不稳定平衡之上或之下,也就是分离矩阵。
,可能接近高平衡或低平衡,取决于起始位置是否在不稳定平衡之上或之下,也就是分离矩阵。
图3.的平衡
 为(16)式。策划反对
为(16)式。策划反对 为
为 .的点
.的点 而且
而且 在上分隔间隔坐标轴上,系统有三个非零平衡。
在上分隔间隔坐标轴上,系统有三个非零平衡。
硬失稳 假设参数
 开始时数值较低,随着森林的生长逐渐增加。事实证明不会随着森林的生长而改变;因此,图3适用。因为成正比
开始时数值较低,随着森林的生长逐渐增加。事实证明不会随着森林的生长而改变;因此,图3适用。因为成正比 ,将会增加。起初,(当
,将会增加。起初,(当 )时,虫的数目将维持在较低水平,因为唯一稳定的均衡是较低的。即使增加之外时,虫的数量将保持较低,因为它们位于不稳定平衡之下,而不稳定平衡决定了低平衡的吸引范围。低平衡的稳定性变得岌岌可危方法,因为吸引力的范围缩小了。最后,在
)时,虫的数目将维持在较低水平,因为唯一稳定的均衡是较低的。即使增加之外时,虫的数量将保持较低,因为它们位于不稳定平衡之下,而不稳定平衡决定了低平衡的吸引范围。低平衡的稳定性变得岌岌可危方法,因为吸引力的范围缩小了。最后,在 时,较低的两个平衡态消失,芽虫密度跳升至较高值:爆发发生。这种吸引状态的突然变化被称为“硬失稳”。它应该与系统显示的软稳定性损失(Eq。9).就芽虫而言,一旦密度达到了较高的平衡,就没有容易的方法将其降低到较低的平衡。如果变量减少低于时,虫体保持高度平衡。作为是进一步减少,有第二个硬失稳作为下面的下降.在这种情况下,有一个向下的跳跃到低平衡,而不是相反再次增加。
时,较低的两个平衡态消失,芽虫密度跳升至较高值:爆发发生。这种吸引状态的突然变化被称为“硬失稳”。它应该与系统显示的软稳定性损失(Eq。9).就芽虫而言,一旦密度达到了较高的平衡,就没有容易的方法将其降低到较低的平衡。如果变量减少低于时,虫体保持高度平衡。作为是进一步减少,有第二个硬失稳作为下面的下降.在这种情况下,有一个向下的跳跃到低平衡,而不是相反再次增加。
磁滞和周期 如果我们现在把树的动态和虫的动态联系起来,一个新的现象就出现了。如果系统从低叶密度和低芽虫数量开始,叶密度会慢慢增加,直到超过
.此时,将发生爆发,如前面所示。大量的芽虫数量最终会导致树木死亡,所以开始减少时,budworm有一个爆发。尽管如此,百虫的数量仍然很高下降,因为芽虫密度在隔膜上方。作为继续下降到时,芽虫密度下降缓慢,后又跳至较低值减少低于.整个系统的不同路径是递增和递减的构成了“滞后效应”。芽虫和森林动态的结合产生了长周期的稳定循环。这种通过快速转换和缓慢变化的交替来维持的稳定循环被称为“松弛振荡”。它们在许多物理、化学和生理系统中都很常见(Edelstein-Keshet 1988)。
干扰和弹性 如果管理的目标是保持budworm的数量和叶子的损害低,失去稳定性
增加之外可能被认为是丧失了韧性。该模型表明,在较低的稳定平衡附近的小扰动如果接近不稳定支路,可能会表现出较长的返回时间。这样长的返回时间可能是一个有用的诊断指标。然而,由于随着树木的生长,稳定的丧失伴随着虫的爆发似乎不可避免。我们可以从不同的角度来看待周期性的虫灾,将其作为不时更新森林的稳定系统的一部分。实际上,类似于竹林系统的系统经常以稳定振荡的形式出现。这种振荡器的优点是,在各种各样的干扰下,它们继续以相同的频率和振幅振荡或多或少。因此,生理振荡对于维持有机体的完整性是很重要的,这是另一种弹性。根据这一观点,从长远来看,试图阻止振荡可能会导致灾难性的崩溃。为了提高自然系统的生产力而进行的人为干预会遭遇类似的命运吗?
湖动力学 Carpenter等人(1996)讨论了这些想法在湖泊生态系统中的适用性。他们将湖泊动力学描述为“正常”或“病态”。正常的湖泊有大量的狩猎鱼,有效地捕食浮游植物,藻华发生率低。当受到诸如磷脉冲之类的干扰时,正常的系统保持其完整性,因为磷会迅速进入较高的营养水平(Carpenter和Kitchell 1993年),腐殖质会限制藻类的生长(Jones 1992年)。然而,过度的磷酸盐负荷、大型植物的移除、过度捕捞、湿地和河岸植被的移除可能会导致野生鱼很少、放牧较少、大型植物没有、广泛而频繁的藻华(Harper 1992)的病态状态。这可能是一个快速的转变,而且不容易逆转(国家研究理事会1992年)。
这种情况似乎符合硬失稳的定义,因为变化迅速而巨大,有时即使磷负荷减少也不会逆转(国家研究理事会1992年)。有人可能会说,正常的湖泊是有弹性的,因为它在扰动下保持其完整性,但弹性会随着磷酸盐负荷和其他应力的增加而丧失。如果可以确定磷酸盐的临界水平和其他环境变量,我们可能会尝试根据与临界水平的差异来衡量弹性(Vollenweider 1976)。也许湖泊生态系统是否符合我们的定义的问题可以通过对长期数据的统计分析来回答。
波罗的海 Jansson和Velner(1995)描述波罗的海系统的术语显示出与湖泊系统的许多相似之处。波罗的海是部分封闭的,因此,水的停留时间约为20年。藻类构成了一个多样化的食物网的基础,较高的营养水平被具有商业价值的物种占据,如鲱鱼、比目鱼、梭子鱼和鲈鱼。由于缺乏来自北海的流入,可能会在很长一段时间内出现较弱的水柱垂直混合。在这期间,更深的地方氧气含量可能很低,硫细菌可能占主导地位。后者将大量的磷放入溶液中,然后向上上升,导致浮游生物大量繁殖。
在历史上,波罗的海经历了几次长时间的缺氧,但这一系统并没有被永久改变。自工业革命以来,波罗的海的磷含量一直在增加,有迹象表明其结构已转变为以碎屑为基础的系统。这将意味着更浑浊的水和主要由活动迟缓的鱼类组成的鱼类群落,如鲷鱼、蟑螂和红鳍鱼,它们的价值比之前列出的要低得多。有一种可能性是,波罗的海可能达到一个点,即使减少磷酸盐的输入也不能使系统恢复到较早较理想的状态。这种事件的转变将相当于稳定的严重丧失,类似于湖泊生态系统的行为。不幸的是,波罗的海航运系统没有复制;因此,我们只有类比来指导行动。有目的的证明波罗的海实际上能够发生与严重失稳相对应的突然变化的实验是不可想象的。然而,它可能是由于人为的疏忽而发生的。波罗的海早期从缺氧期恢复的能力可能与韧性相对应,但我们不能确定这种韧性是否正在丧失。
北方森林的 Carpenter et al.(1996)将针叶林描述为一个物种相对较少、相互作用和动态复杂的系统。在高地地区,以白杨和桦树为主的森林与以云杉和冷杉为主的森林交替生长。驼鹿在20-40年的时间里浏览可以把白杨林变成针叶树占主导地位的林场。随着针叶林的成熟,它们变得越来越有利于云杉芽虫的繁殖。最终,爆发发生,部分系统转化为早期演替白杨。芽虫的爆发对应着稳定的硬丧失,合并的高地系统经历了类似于前面描述的稳定的长周期振荡。
由于高地地区经历了这种振荡,山谷底部在被洪水淹没的平原和潮湿的草地之间交替。洪水泛滥的状态是由海狸维持的,它们砍伐溪流边的白杨作为食物,并在溪流上筑坝形成池塘。当白杨的供应不足时,海狸就会放弃它们的水坝,水坝就会破裂,池塘很快就会被草地取代。这种相对迅速的变化是白杨供应减少的结果,可以被认为是一种稳定的硬丧失。由于海狸和白杨的相互作用,高地和低地的循环往往相互交织。由于被云杉芽虫杀死的针叶树提供了充足的燃料,火灾也在大范围内同步循环中发挥了作用。
尽管这个系统经历了很大的变化,有时是非常快的,但它可能被认为是有弹性的,在许多世纪中保持其特性。任何地点的情况都可能突然变化,但整个系统通常是在周期的不同阶段由多个补丁拼接而成。当作为一个整体考虑时,它保持了相当大的多样性。
想象一下,从高水平的草和低水平的木本植被开始。在放养水平较低的情况下,与非放牧系统只有很小的区别:如果系统一开始以草为主,那么草将继续占主导地位。随着蓄水量的增加,竞争可能有利于木本植物。最终,可能会有草的崩溃,木本植物将占主导地位。因此,放牧的效果是将系统从草占主导地位的状态转移到木本植被占主导地位的状态。即使在放牧压力减轻的情况下,植物的组成也可能没有什么变化,因为当木本植物占主导地位时,木本植物比草具有优势。放牧的作用是使该系统进入木本植被对非放牧系统的吸引范围。
如果绘制牧草密度与放养水平的关系图,这种行为可能会显得难以解释:随着放牧的增加,牧草水平下降,但当放牧恢复到以前的水平时,却不会恢复到以前的水平。如果我们认识到草的密度不仅取决于饲养水平,而且取决于与木本植物的竞争,那么这个明显的悖论就解决了。这些现象可以通过修改Lotka-Volterra竞争模型来说明。
数学模型 让
 表示草的密度,并让
表示草的密度,并让 表示木本植被密度。草密度的变化率假设为
表示木本植被密度。草密度的变化率假设为
在哪里
 是增长率,还是
是增长率,还是 而且
而且 是竞争系数。的参数
是竞争系数。的参数 由牛的放养率决定。假定木本植被的增长率为
由牛的放养率决定。假定木本植被的增长率为
在哪里
 是增长率,
是增长率, 而且
而且 竞争系数,和
竞争系数,和 是一个源项。在下面的插图中,参数被选择为:
是一个源项。在下面的插图中,参数被选择为:
轻放牧的情况对应于
= 1/10。
为了理解该系统的行为,在模型中绘制一些曲线是有帮助的 面,如图4.在相平面上,系统的变化方向和速度由矢量给出
面,如图4.在相平面上,系统的变化方向和速度由矢量给出 .这个向量垂直于曲线
.这个向量垂直于曲线 (零等斜线),在曲线上是水平的
(零等斜线),在曲线上是水平的 (零等斜线)。可以看出,,要么
(零等斜线)。可以看出,,要么 ,或
,或
这个轨迹是一条直线,它向左移动增加。零-等斜线是根据(Eq。18).其中一条渐近线是-轴,轨迹通过该点 ,贴上“w”这个轨迹与存货参数无关.图4详细展示了一个系统如何根据启动条件接近多个稳态。虽然这个系统比(要复杂得多)Eq。9),则其定性行为相同
,贴上“w”这个轨迹与存货参数无关.图4详细展示了一个系统如何根据启动条件接近多个稳态。虽然这个系统比(要复杂得多)Eq。9),则其定性行为相同 .这说明了非常简单的一维模型是如何成为有价值的启发式指南的。
.这说明了非常简单的一维模型是如何成为有价值的启发式指南的。
图4.相平面为草与树,按(17)式。,Eq(18)。,Eq。(19).零
 而且
而且 等斜线用虚线标出。这些是曲线
等斜线用虚线标出。这些是曲线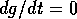 或
或 ,分别。平衡点是等斜线相交的点。稳定平衡分别标记为“G”和“W”。不稳定平衡称为s。进入S的轨迹形成分隔线:它们用虚线标出。分离轴右侧的轨迹最终到达G,左侧的轨迹最终到达W。
,分别。平衡点是等斜线相交的点。稳定平衡分别标记为“G”和“W”。不稳定平衡称为s。进入S的轨迹形成分隔线:它们用虚线标出。分离轴右侧的轨迹最终到达G,左侧的轨迹最终到达W。
我们现在谈谈增加存货的影响。增加的影响
是移空值吗-等斜线向下向左。因此,点S和G会沿着空等斜线。因为分离线经过S点,G的引力域必然缩小,而W的引力域则会扩大。定性上,表示图4仍然成立。对于更高的值, ,点S和G重合。的值对应的根S和G如所示图5.的对应值可以画一个类似的图来表示.这是一个类似于图3.如果载畜率变化缓慢,我们可以期望草密度由中上部曲线给出图5.然而,如果
,点S和G重合。的值对应的根S和G如所示图5.的对应值可以画一个类似的图来表示.这是一个类似于图3.如果载畜率变化缓慢,我们可以期望草密度由中上部曲线给出图5.然而,如果 ,草密度必须崩溃,因为在草密度非零的情况下不存在稳定的平衡。这是一种硬失稳,两者确实有惊人的相似之处无花果。而且3..
,草密度必须崩溃,因为在草密度非零的情况下不存在稳定的平衡。这是一种硬失稳,两者确实有惊人的相似之处无花果。而且3..
图5.显示草均衡作为放牧参数的函数的分岔图
 对于式(17)-式(19)。如果
对于式(17)-式(19)。如果 ,有三个根。最小的分支对应于点W
,有三个根。最小的分支对应于点W ,这总是一个稳定的平衡。中间的分支对应于点S的轨迹为各不相同。这些点是不稳定均衡,所以用虚线表示。最高的分支是点G的轨迹各不相同。这个点总是稳定的。
,这总是一个稳定的平衡。中间的分支对应于点S的轨迹为各不相同。这些点是不稳定均衡,所以用虚线表示。最高的分支是点G的轨迹各不相同。这个点总是稳定的。
的值
的定性形式图6适用。唯一的平衡点是W,所有的轨迹都接近W, W的引力域是整个象限 而且
而且 .
.
图6.系统(17)-(19)的相平面为草和树,其中
 .在这种情况下,系统只有一个稳定平衡.所有的轨迹都接近这个点。
.在这种情况下,系统只有一个稳定平衡.所有的轨迹都接近这个点。
我们可以想象系统从载畜率开始
 .根据图5,不稳定平衡S和稳定平衡W非常接近。因此,分隔线在图中类似于图4在最左上角:几乎任何初始组合而且会不会导致高密度,由上面的分支给出图5,密度相应较小.现在,如果是慢慢增加,密度的会沿着上分支向下移动直到分叉点吗
.根据图5,不稳定平衡S和稳定平衡W非常接近。因此,分隔线在图中类似于图4在最左上角:几乎任何初始组合而且会不会导致高密度,由上面的分支给出图5,密度相应较小.现在,如果是慢慢增加,密度的会沿着上分支向下移动直到分叉点吗 是达到了。超过这一点,草的密度就会崩溃,木本植物就会占主导地位。即使放牧压力被撤消(),草无法恢复,因为它将在分离体的左边。这就是滞后效应。
是达到了。超过这一点,草的密度就会崩溃,木本植物就会占主导地位。即使放牧压力被撤消(),草无法恢复,因为它将在分离体的左边。这就是滞后效应。
从长远来看,短时间的放牧可能会导致草的迅速崩溃,随后是缓慢的恢复。从短时间尺度上看,这似乎是一个具有高木本植被的平衡,但从长时间尺度上看,这对应着该系统在该区域花费了很长时间。这种定性行为类似于“可兴奋系统”。根据霍奇金和赫胥黎的理论,这种系统被称为神经冲动的模型,并经菲茨休修正。该系统有一个单一的稳定平衡,但当在适当的方向受到干扰时,它可能会经历一个非常大的偏移(神经元放电),然后是一个很长的恢复(不应)期。详细内容见edelstein - kesheet(1988)。
火对动力学的影响 正确地模拟老化是复杂的,但就目前的目的而言,找到一个具有所需定性行为的简单系统就足够了。我们并不认为下面的模型是真实动态的准确表示。我们必须把多余的草记录下来,因为它们可能会成为生火的燃料。因此,我们让草生产总量由
 ,在那里
,在那里
在哪里
 是一个源项,和是可用于放牧的草的生产速率。没有被放牧的牛消耗掉的草是潜在的火灾燃料,用
是一个源项,和是可用于放牧的草的生产速率。没有被放牧的牛消耗掉的草是潜在的火灾燃料,用 ,在那里
,在那里
的参数
确定牛所消耗的草的比例。现在(Eq.17)由
的动力
会受到火灾的影响,树木的年龄影响它们对火灾的易感性。让一个新变量 表示木本植物密度与木本植物平均树龄的乘积。第一个近似的结果是
表示木本植物密度与木本植物平均树龄的乘积。第一个近似的结果是 但这种关系忽略了火灾对平均年龄的影响。的动力而且是由
但这种关系忽略了火灾对平均年龄的影响。的动力而且是由
哪里有火灾风险
 定义如下:让火势
定义如下:让火势 与可用燃料成正比:
与可用燃料成正比:
我们假定火灾风险
是由
的参数
 火灾风险是其最大风险的一半的年龄和参数
火灾风险是其最大风险的一半的年龄和参数 决定了火灾风险随年龄增加的锐度。我们使用
决定了火灾风险随年龄增加的锐度。我们使用 给一个大幅度的增加,和
给一个大幅度的增加,和 .其余参数选择为
.其余参数选择为
的载畜率
可选择作为参数或控制变量。在下面,我们选择 .
.
因为在这个系统中有三个状态变量,所以不能画出有意义的相位图。然而,如果年龄变量是固定的,我们可以获得动态的印象。图7说明一个低的值使系统达到一个高的平衡.另一方面,越高的值导致一个低得多的平衡,如图8.现在,如果随着时间的增加,系统会先有高,那么低因为它年龄。火灾降低了树木的平均年龄和密度。因此,系统重置到类似于中所示的状态图7后起火。骑自行车的加上整个系统的时间,加上变化,表现在图9.
图7.(21)-(28)系统中草和树的相平面,忽略了年龄变量的动态变化
 的值较低.只有一个高值的平衡和较低的值,是稳定的。所有的轨迹都接近这个平衡。
的值较低.只有一个高值的平衡和较低的值,是稳定的。所有的轨迹都接近这个平衡。
图8.(21)-(28)系统中草和树的相平面,忽略了年龄变量的动态变化
的高值.的单一平衡值较低和一个中等高的值.
图9.树密度(W) vs.时间(T为系统(21)-(28),包括年龄变量的动态。对于草密度也可以画出类似的图,当木本植被较低时,草密度往往较高,反之亦然。
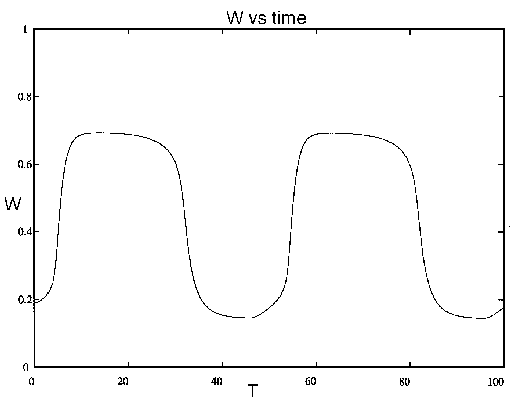
如果观察这个系统5-20年的时间,就会发现,由于竞争和放牧的结合,木本植物最终将主宰草类植物。然而,在接下来的30年里,火灾和树木老化的影响导致树木倒塌,为下一个周期让路。这个例子说明了我们观察系统的时间尺度如何对我们对其行为的分类产生决定性的影响。不幸的是,我们能够观察系统的尺度往往比它表现其特征行为的尺度短得多。
在这里,我们从简单的概念模型,如木筏类比,到简单而抽象的数学模型,再到生态系统类比,最后到一个相当详细的草原系统模型。在任何情况下,这些类比都不能被认为是完整的,我们对系统的知识也不够详细,不足以支持一个具有统计理由的成熟模型。也许有人会得出这样的结论:所有这些都只是猜测,不值得认真关注。另一方面,谨慎的决策要求我们考虑到对我们的行为的反应的各种似是而非的假设。这里举的例子并没有鼓励人们自满,认为自然系统有能力以我们过去所享受的奢侈方式支持我们和我们的习惯。如果我们拒绝思考那些隐约可见的可能性,我们就可能失去了解世界并相应地调整行为的机会。如果我们在选择行动时坚持把最简单、最方便的假设放在首位,我们就会冒着让懒惰和骄傲给我们带来灾难的风险。
欢迎对本文进行回复。如果被接受发表,您的回复将被超链接到文章。要提交评论,请关注这个链接.要阅读已接受的评论,请关注这个链接.
该研究是由瑞典皇家科学院下属的Beijer研究所支持的“弹性网络规划研讨会”的成果。
- 阿罗,K, B.博林,R.科斯坦扎,P.达斯古普塔,C.福尔克,C. S.霍林,b - o。詹森,莱文,k。Mäler, C. Perrings和D. Pimentel。1995.经济增长、承载能力和环境。科学 268: 520 - 521。
- 卡朋特,s.r., b.o.简森,D.路德维希,J.帕斯特,G.彼得森,B.沃克。1996.系统总弹性的比较分析。4月10日至16日北京理工大学恢复力网络规划研讨会的说明.
- 卡朋特,S. R.和J. F.基切尔。1993.湖泊中的营养级联。剑桥大学出版社,英国剑桥。
- 卡朋特,s。R。和p。R。莱维特。1991.由营养级联引起的古生态记录的时间变化。生态 72(1): 277 - 285。
- 德安东尼奥,c·M·维图塞克。1992.外来草的生物入侵,草火循环和全球变化。生态学与系统学年评 23: 63 - 87。
- 完成,t·J。1992.珊瑚礁群落的相移及其生态意义。Hydrobiologia 247: 121 - 132。
- 都柏林,H. T., A. R. E.辛克莱,J.麦克格莱德。1990.大象和火灾是塞伦盖蒂-马拉林地多重稳定状态的原因。动物生态学杂志 59: 1147 - 1164。
- “L。1988.生物学中的数学模型。兰登书屋,纽约,纽约,美国
- 埃斯蒂斯,j。A。和d。达金斯。1995.阿拉斯加海獭和海带林:群落生态范式的共性与变异。生态专著 65(1): 75 - 100。
- 古肯海默,J.和P.霍姆斯。1983.非线性振荡,动力系统,和向量场的分岔。Springer-Verlag,纽约,纽约,美国。
- 哈珀,D。1992.富营养化的相似性。查普曼和霍尔,伦敦,英国。
- 温和,c . S。1973.生态系统的恢复力和稳定性。生态学与系统学年评 4: 1。
- 休斯,t . P。1994.灾难,相位变化,以及加勒比海珊瑚礁的大规模退化。科学 265: 1547 - 1551。
- 简颂B.-O。,H. Velner.1995.波罗的海:充满惊奇的海洋。292 - 372页在l·h·甘德森,c·s·霍林和s·s·莱特,编辑。生态系统和制度更新的障碍和桥梁。哥伦比亚大学出版社,美国纽约。
- 琼斯,r . I。1992.腐殖质物质对湖泊浮游食物链的影响。Hydrobiologia 229: 73 - 91。
- 路德维希,D.琼斯和C. S.霍林。1978.云杉芽虫与森林昆虫爆发系统的定性分析。动物生态学杂志 47: 315 - 332。
- 麦克拉纳汉,T. R., A. T. Kamukuru, N. A. Muthiga, M. Gilagabher Yebio, D. Obura。1996.海胆减少对藻类、珊瑚和鱼类数量的影响。保护生物学 10(1): 136 - 154。
- 国家研究委员会。1992.恢复水生生态系统。国家学院出版社,华盛顿特区,美国。
- •奥德姆,e . P。1993.生态和我们濒临灭绝的生命维持系统。第二版。西诺尔,桑德兰,马萨诸塞州,美国。
- 皮姆,s . L。1991.自然的平衡?芝加哥大学出版社,美国伊利诺斯州芝加哥。
- 舍弗,M. S. H.霍珀,M. l。梅杰,B.莫斯,E.杰普森。1993.浅湖的可选平衡。树 8(8): 275 - 279。
- 辛德勒,d . W。1990.整个湖泊的实验扰动,作为生态系统结构和功能假说的检验。Oikos 57: 25-41。
- Vollenwieder, r。1976.湖泊富营养化磷临界负荷水平的研究进展。意大利idro生物学研究所纪念 33: 53 - 83。
- 沃克,B. H.路德维希,C. S.霍林,R. M.彼得曼。1981.半干旱草原放牧系统的稳定性。《生态学杂志》 69: 473 - 498。
- 兹莫夫,s.a., v.i.查普林,a.p. Oreshko, F. S.查宾,III, J. F.雷诺兹和M. C.查宾。1995.草原-苔原过渡:更新世末期草食动物驱动的生物群转移。美国博物学家 146(5): 765 - 794。
回归时间和弹性区分靠近稳定平衡的行为和靠近吸引域边界的行为是很重要的,这是一个不稳定的平衡或分离边界。正如第2节所讨论的,与弹性损失相关的长返回时间是由靠近的缓慢动力引起的不稳定平衡,不是靠慢动力学靠近的稳定的平衡点。不幸的是,对于弹性的定义存在两种相互冲突的定义,并因此混淆了弹性与回归时间之间的联系。
皮姆(1991:13)将弹性定义为“一个变量从平衡中被转移到平衡中的速度。”弹性可以通过返回时间来估计,即位移衰减到其初始值的某个特定部分所需的时间。皮姆(1991:33)通过方程描述了回归平衡
在哪里 人口密度与时间有关吗
人口密度与时间有关吗 ,
, 是初始人口密度,和
是初始人口密度,和 是平衡密度。的微分方程对应于这个公式的是
是平衡密度。的微分方程对应于这个公式的是
可以给出一个离散时间的类似模型,但这不会改变下面的论证。如果我们测量位移通过 ,然后满足
,然后满足
这相当于Eq。(.)如果 取而代之的是.严格地说,皮姆的定义依赖于这种简单性,因为所需的时间数量衰减到其初始值的某一特定部分只有当模型(.)使用。事实上,如果初始位移是
取而代之的是.严格地说,皮姆的定义依赖于这种简单性,因为所需的时间数量衰减到其初始值的某一特定部分只有当模型(.)使用。事实上,如果初始位移是 分数是
分数是 ,然后(.)意味着
,然后(.)意味着
由此我们得出回归时间 是由
是由
显著的特征是没有出现在这个公式中。这只是这个模型的一个特性,如下所示。在更一般的情况下,这样的结果只能在极限为时成立 .这样的结果被称为“局部的”。正如第2节所指出的,一个常见的错误是将局部结果外推到全局结果。在目前的情况下,它相当于用线性近似代替一个复杂的函数。这样的近似当然很容易处理,但它们可能会错过动力学的基本特征。事实上,未能认清局部稳定和全球稳定之间的区别,可能会导致人们对干预自然系统可能产生的后果持毫无根据的乐观态度。如果我们认为对小扰动的稳定性必然意味着对大扰动的稳定性,那么就不需要预防。
.这样的结果被称为“局部的”。正如第2节所指出的,一个常见的错误是将局部结果外推到全局结果。在目前的情况下,它相当于用线性近似代替一个复杂的函数。这样的近似当然很容易处理,但它们可能会错过动力学的基本特征。事实上,未能认清局部稳定和全球稳定之间的区别,可能会导致人们对干预自然系统可能产生的后果持毫无根据的乐观态度。如果我们认为对小扰动的稳定性必然意味着对大扰动的稳定性,那么就不需要预防。
为了区分平衡点附近的行为 从接近不稳定平衡的行为来看,我们必须使用一个参数多于(a).我们设置
从接近不稳定平衡的行为来看,我们必须使用一个参数多于(a).我们设置
这种特殊的形式引出了一个特别简单的返回时间方程:到达一个位置的时间 从是由
从是由
还有表格 被选中是为了
被选中是为了
可以用代数方法验证。针对(A.7)而且(如),
现在,如果我们替换通过 ,(A.9)就变成了
,(A.9)就变成了
在哪里
如果最后两项在(A.10),这个结果与皮姆的假设是一致的吗(.).我们更复杂的动力学假设(要求寄出)是皮姆假设的类似物如果存在三个均衡。在什么情况下(A.10)意味着更长的返回时间?第一项,对应于皮姆的模型,意味着一个较长的回报时间,如果比率 是小的,如果
是小的,如果 很小。在皮姆的讨论中,
很小。在皮姆的讨论中, 是描述系统探测或观测结果的参数。通常,是固定的,而返回时间提供了一个估计为.
是描述系统探测或观测结果的参数。通常,是固定的,而返回时间提供了一个估计为.
第二项(A.10)意味着返回时间很长,如果 小或
小或 很小。我们之前的讨论涉及到一个可能的变量
很小。我们之前的讨论涉及到一个可能的变量 以及可能使系统接近不稳定平衡的扰动。对应于附近
以及可能使系统接近不稳定平衡的扰动。对应于附近 ,或附近
,或附近 .在这种情况下,将是大的,即使参数很大。也就是说,返回时间可能很长,即使对于接近稳定平衡时显示非常快速返回的系统也是如此。根据这个观点,长返回时间可能诊断为小
.在这种情况下,将是大的,即使参数很大。也就是说,返回时间可能很长,即使对于接近稳定平衡时显示非常快速返回的系统也是如此。根据这个观点,长返回时间可能诊断为小 或者是大到足以使系统接近不稳定平衡的扰动。它们也可能对应于来自不稳定平衡的弱斥力,即,小.如果一个扰动使系统超出了不稳定的平衡,就没有任何回报。
或者是大到足以使系统接近不稳定平衡的扰动。它们也可能对应于来自不稳定平衡的弱斥力,即,小.如果一个扰动使系统超出了不稳定的平衡,就没有任何回报。
总而言之,根据Pimm(1991)和我们的观点,较长的返回时间可能被诊断为丧失弹性,但这两种情况下术语的含义是完全不同的。皮姆关注的是接近稳定平衡的行为。在这种情况下,一个给定的位移从平衡的返回时间较长表明系数较小或者,等价地,一个小的导数 .我们关注的是具有两个或三个平衡的系统的行为,其中一个是稳定的。弹性描述了系统回归其稳定平衡的趋势。较长的返回时间是由于扰动使系统接近不稳定平衡,或可能是来自不稳定平衡的弱斥力。
.我们关注的是具有两个或三个平衡的系统的行为,其中一个是稳定的。弹性描述了系统回归其稳定平衡的趋势。较长的返回时间是由于扰动使系统接近不稳定平衡,或可能是来自不稳定平衡的弱斥力。
记者的地址:
唐纳德·路德维希
数学系
英属哥伦比亚大学
加拿大不列颠哥伦比亚省温哥华V6T 1Z2
电话:604-541-9409
传真:604-822-6074
ludwig@math.ubc.ca
合著者的地址:
布莱恩•沃克
CSIRO野生动物与生态部
公署箱84
Lynham, ACT 2602,澳大利亚
电话:6-242-1742
传真:6-241-1742
brian.walker@dwe.csiro.au
克劳福德s温和
佛罗里达大学
动物系,邮箱118525
美国佛罗里达州盖恩斯维尔32611
电话:352-392-6914
传真:352-392-3704
holling@zoo.ufl.edu
*这篇文章的版权于2000年1月1日从美国生态协会转让给韧性联盟。
![]()