|
版权所有©1997韧性联盟*
沃克,B.和W.斯特芬,1997。全球变化对自然和管理陆地生态系统的影响概述。保育生态[网上]1(2): 2。可从互联网获得。URL: http://www.consecol.org/vol1/iss2/art2/本文的一个版本,其中的文本、图表、表格和附录是单独的文件,可以通过以下命令找到链接.
合成全球变化对自然和管理陆地生态系统的影响概述*布莱恩•沃克1而且将史蒂芬1
1CSIRO野生动物与生态和GCTE核心项目办公室
全球变化是土地利用、大气成分、生物多样性和气候变化的个体和相互作用的净效应。IGBP(国际地圈-生物圈计划)的全球变化和陆地生态系统项目过去六年的活动综合研究了作为生态系统响应和与全球变化共存的全球变化效应。生态系统响应是根据功能和植被组成/结构的变化来考虑的。高浓度CO的现场试验2对地上生物量的影响平均表现为正影响,范围为-20%至+80%。CO的一些早期预测2影响(C3.与C4植物、N固定物、凋落物中的C:N)一般不受支持,有必要考虑CO变化的相互作用2温度和氮。涉及瞬时变化的动态全球植被模型表明,生物群落不会作为完整的实体发生变化。下个世纪,植被,特别是高纬度地区的植被可能发生重大变化,而扰动体系的变化将是最重要的。根据对土地利用、植被结构和生态系统生理学变化的预测,下个世纪陆地生物圈可能会成为碳的来源而不是碳汇。
由于土地利用的变化,21世纪的陆地生物圈可能会在物种丰富度上进一步减少,并进行大规模的重组。更多的自然生态系统将处于早期演替状态或转变为生产系统。生物圈通常会杂草丛生,结构更简单,生态复杂的原始生长状态的区域更少。
由于二氧化碳的排放,温带作物的产量可能会略有增加2增加(平均田间条件下小麦产量为5-7%),但热带地区某些地区的作物产量可能下降。土地利用变化对牧场和牧场的影响最大;由于需要每年增加2%的作物产量来满足不断增长的人口,农田将会越来越多地入侵牧场。
提交日期:1997年10月24日。录用日期:1997年11月19日。
关键词:农业;生物群落;二氧化碳;气候;动力学;生态系统功能;全球变化;影响;土地利用;陆地生物圈; vegetation changes.
评估世界自然和管理的陆地生态系统的未来的出发点是认识到全球变化远不止是气候变化。beplay竞技它是土地利用、大气成分、生物多样性、而且beplay竞技气候变化。关于这四大全球变化驱动因素的现状,我们注意到:
-
•大气成分正在变化。CO的增加2和其他温室气体是无可争议的,除了氟氯化碳,它们至少在未来几十年还会继续上升。
-
-
•土地利用正在发生变化。这包括从一种土地覆盖类型向另一种土地覆盖类型的转换,以及管理实践的改变(如施肥、从耕作转变为免耕等)。它还包括地表水可用性和河流流量的变化。
-
-
•世界大部分地区的生物多样性正在下降。
-
-
•气候可能正在变化。政府间气候变化专门委员会(IPCC) 1995年的评估(IPCbeplay竞技C WGI 1995)得出结论,人类引起的世界气候变化是可探测的。
在这四个驱动因素的背后,是人类人口无可争议的巨大变化(图1),以人口、人口流动、自然资源的人均消耗量和获取资源的技术能力的增长来衡量。
图1所示。人口增长。
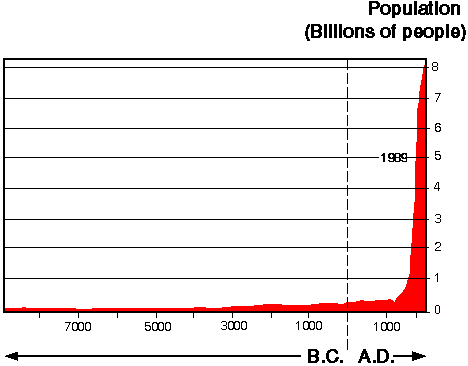
这些统计数据广为人知,但认识到其重要性并强调以下几点是很重要的。尽管联合国最近的估计表明,到下个世纪中叶以后,人口增长率将略有下降,但在此之前,人口将继续以目前的速度增长。所有评估都认为,到2050年,人口增长将不会放缓。联合国估计2050年的人口中位数(最有可能的)是94亿,比上世纪90年代增加了约80 x 106每年的人口。自1900年以来,人口增长了三倍,与此同时,世界经济增长了20倍,化石燃料消耗增加了30倍,工业生产增加了50倍,当人们认识到这一点时,人口变化的影响就变得更加清晰了。在未来的50年里,人口数量将再增加两倍,世界经济将增长5到10倍。
由于人口增长不可阻挡,农作物产量必须每年持续增长2%;在下个世纪中叶之前,它们必须增长250%。经济发展和同时的技术进步相结合,将导致自然资源的人均消费量增加。这些发展的生态后果将不可避免地包括土地利用做法的进一步集约化和自然植被进一步转为耕地,大部分是在目前不适合种植的地区。
回到我们的开篇,大气成分和气候的变化将影响土地利用。这些土地利用的变化将直接反馈给大气成分和气候;所有这些变化(统称为全球变化)的净影响将是它们各自的和相互作用的综合影响。
本文基于国际地圈-生物圈计划(IGBP)全球变化和陆地生态系统(GCTE)项目前六年的综合研究。该综合研究由包括GCTE科学指导委员会(SSC)在内的30名科学家进行,并基于涉及全球44个国家的1000名科学家和技术人员的大量研究(GCTE研究计划)。综合(Walker等人,在新闻),预计于一九九八年年初出版。该论文的格式遵循了综合执行摘要的第一作者在1997年8月美国生态学会年会上的报告。我们在这里将综合的主要发现分为三个部分:生态系统响应、陆地碳循环和与全球变化共存。
生态系统的响应可以从两个方面来考虑:生态系统功能和植被变化。第一个研究涉及陆地生态系统与全球变化的相互作用,前提是给定一个生态系统的任何特定状态。第二部分涉及全球变化导致的生态系统状态(即植被的结构和组成)的变化。
生态系统功能许多关于生态系统生理学的初步工作都集中在CO升高对生态系统水平的影响上2以及它与其他因素的相互作用。新出现的认识可以总结如下:
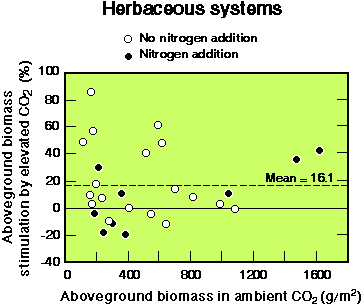
- •大多数生态系统暴露于双重CO2生长旺季净碳吸收量高于环境CO2浓度。对于草地来说,在所有实验中,地上生产力平均增加了16%,尽管个体草地群落的反应差异很大,有些是负面的(见图2).群落响应的变化反映了其组成物种的变化、物种间的相互作用以及CO的高度相互作用性质2与其他环境因素的反应,如水,养分有效性和温度。
图2所示。高浓度CO对草本植物地上生物量的刺激2,加氮和不加氮。所示的平均值适用于所有点,无论是否添加氮。
-
-
•苔原和高山草原等寒冷生态系统对CO升高的反应最小2,在某些情况下没有生长反应,并在几年后完全适应旺季气体交换。幼树的快速生长并不能说明森林作为一个整体是否会吸收更多的碳。
-
-
•两个关于CO升高反应性的早期预测2(1) C4物种的反应小于C3.物种;(2)具有固氮共生体的物种生物量响应更大。总的来说,这些预测都没有在生态系统研究中得到一致的证实。
-
-
•与之前的预测相反,在高浓度CO下生长的植物产生的垃圾2不一定分解得更慢。高浓度CO下植物凋落物中C:N的比值2一般不高于在CO环境中生长的植物2,尽管物种之间有很大的差异。
-
-
•高有限公司2一般增加光合产物分配到根,这增加了地下碳汇的容量和/或活性。模型表明,地下碳汇容量的增加可能导致长期土壤碳固存量的增加,尽管仍缺乏强有力的经验证据。
-
-
暴露于高浓度CO的草本植物2气孔导度降低,通常导致土壤水分流失减少。这种水可用性的增加是有限水草地系统净碳吸收量增加的主要驱动因素。
-
-
•暴露于高浓度CO的树苗气孔导度也会降低2,但根据目前的实验数据集,这似乎不是成熟树木(森林)的情况。在大树中,气孔导率的控制作用明显较弱。
-
-
•温度的间接影响将比直接影响更显著。由于热适应,空气温度升高对植物生长的直接影响可能比通常预期的要小。然而,这可能会加速凋落物分解的发展。间接温度效应主要与高纬度永久冻土变暖有关,这可能导致热岩溶扩张,物种组成发生重大变化(向木本灌木而不是苔藓),以及养分有效性增加。
-
-
•在一些森林中,氮沉积与净初级生产力(NPP)的增加有关。然而,从长期来看,持续的氮负荷将导致物种组成的变化,这可能与生态系统层面的碳固存增加有关,也可能与此无关。在许多情况下,持续的氮负荷以及其他相关污染物可能导致土壤酸化,进而导致NPP减少。
-
-
•模型结果表明CO升高的联合效应2土壤温度升高会增加氮矿化和NPP,而土壤温度升高会降低碳储量。
-
-
•对流层臭氧对生态系统NPP有负面影响,但CO升高2是否有可能改善那些在CO升高时气孔导度下降的物种对植物臭氧的伤害2.此外,UV-B辐射的增加也有可能降低NPP。
-
-
•一般来说,只要人类活动对水和养分循环以及扰动机制有直接而重大的影响,这种影响将超过任何直接的CO2对生态系统功能的影响
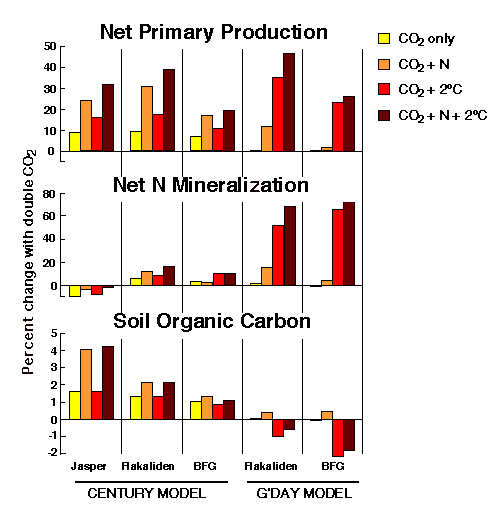
到目前为止,GCTE和相关项目的工作表明,从单一植物的实验推断整个生态系统对全球变化的响应必须非常谨慎(例如,Körner 1995)。同时,公司2不会独立增加,没有其他变化。特别是,温度和大气中氮含量也会发生变化,我们需要了解的是这三种变化的相互作用。一个建模实验(图3)来调查这些交互影响,说明了它们的重要性,也说明了模型差异的重要性。随着研究的进行,其他环境因素的变化将变得明显,需要纳入综合分析。
图3所示。模拟双CO的冲击2,空气温度升高(+2oC)和大气N沉降(+0.25 gx米-2x年-1),利用CENTURY和G’day模型,对三个地点进行了研究:瑞典湿润的挪威云杉地(Flakiliden)、澳大利亚干燥的松林(BFG)和加利福尼亚州的一年生草地(Jasper)。(G’day模型没有在Jasper数据上运行)。
植被变化
第一代区域和全球尺度的植被变化模型基于植被与非生物环境处于平衡状态的假设。这些“平衡”模型很快在广泛的影响研究中得到应用,但这种使用往往会在非生态学家中导致一种误导性的概念,即基于现有生物群落的重新排列的植被变化,导致生物群落从一种平衡分布到另一种平衡分布的急剧转变。
但现实却截然不同。从斑块到全球范围内,对植被组成和结构的影响正在发生,是持续的,可能会加速,并且没有可识别或可预测的终点。从这些植被组成和结构变化的非平衡、瞬态动态中产生的认识包括以下几个重要特征:
-
-
•生物群系不会作为完整的实体发生变化。物种在竞争能力(例如,生长速度)、迁移速度、从干扰中恢复(对干扰的反应)以及其他方面做出不同的反应。因此,新的物种组合将会出现。
-
-
•旧石器研究和模型模拟表明,许多植物物种能够以足够快的速度迁移,以跟上预计的气候变化,但前提是它们能够在连续的、相对不受干扰的自然生态系统中迁移。这强调了自然生态系统碎片化作为全球变化现象的重要后果(Pitelka et al. 1997)。
-
-
•外来物种入侵自然生态系统是一个日益严重的问题。土地利用/覆盖变化的趋势、贸易全球化的加剧和干扰的增加可能会加剧这种情况。
-
-
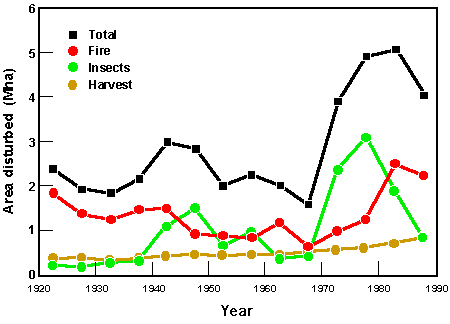
•干扰(例如,火灾、虫害引起的枯死病)似乎在一些地区(例如,北方森林)有所增加,导致更多的生态系统处于早期演代状态(Kurz et al. 1995;看到图4),而在其他区域(例如北欧),管理方面的变化往往会减少早期继承国家的森林面积。
-
-
•由于土壤、土地利用和地形变化的局部影响,在不同的景观中,气候变beplay竞技化对物种组成的影响显著不同。
图4所示。1920 -1988年期间,加拿大北方森林每年受森林火灾、昆虫引起的林分死亡率和砍伐的平均面积(来自Kurz et al. 1995)。
当人们把所有这些因素综合在一起时,关于21世纪植被动态的一些概括开始出现:
-
•由于土地利用的变化,21世纪陆地生物圈的物种丰富度可能会进一步下降,物种组成可能会大幅“重组”,这对生态系统功能的影响尚不可知。
-
-
•随着越来越多的长寿生物(树木)远离最佳环境,并受到土地利用变化带来的越来越大的压力,扰动和枯死病可能会增加。
-
-
•考虑到预计干扰的增加,更多的自然生态系统将处于早期演替状态,或将转变为人类主导的陆地生产系统。这些趋势将导致一个总体上“杂草丛生”的生物圈,结构更简单,生态更复杂的原始生长状态下的系统更少。
陆地生态系统吸收大量CO的潜力2,从而减缓CO的积累2以及减缓气候变化的速度,是关于CO辩论的关键问题beplay竞技2排放控制。目前对全球碳循环的理解,基于已知CO源和汇的预算2为20世纪80年代,显示在表1.
表1。CO的平均年度预算21980-1989年的扰动(来自Schimel 1995年)。碳通量和储层变化用10表示12公斤/年(= Gt/年)。数字来自IPCC(1994年),此外还有来自Schimel的陆地碳汇估计。
| 有限公司2预算 | 1012公斤/年 | 错误* |
| 有限公司2来源 | ||
| 化石燃料燃烧和水泥生产的排放 | 5.5 | 0.5 |
| 热带土地利用变化造成的净排放 | 1.6 | 1.0 |
| 人为排放总量 | 7.1 | 1.1 |
| 有限公司2汇 | ||
| 大气储存 | 3.2 | 0.2 |
| 海洋吸收 | 2.0 | 0.8 |
| 北半球森林再生吸收 | 0.5 | 0.5 |
| 有限公司2受精 | 1.0 | 0.5 |
| N沉积 | 0.6 | 0.3 |
| 剩余(源) | (0.2) | (2.0) |
*误差通过积分累积。
这表明,陆地生物圈在CO的排放和吸收方面基本处于平衡状态2这一结论得到了最近对大气O的测量结果的支持2浓度(例如,Heimann 1997)。估计1.6 x 1012热带地区土地利用变化释放了16亿吨碳,而约2.1 x 1012陆地生态系统通过森林再生、CO2施肥,氮沉降。一些地点已经证明了陆地碳库规模的增加,但尚未在全球范围内或在整个植被扰动周期内得到证实。这种增加很可能稀疏地分布在广泛的生态系统中,因此很难探测到。关键的问题是,陆地生物圈目前是否有能力吸收二氧化碳2可以在将来保持或增加。
预测未来的情况是困难的,因为条款表1不能可靠地预测到未来。与1980年代的扰动预算有关的不确定性已经很大,对未来的具体预测将适用更大的不确定性。但是,可以从预算的角度来评估可能的趋势,它们的相对重要性是增加还是减少,以便可以预测总的趋势。
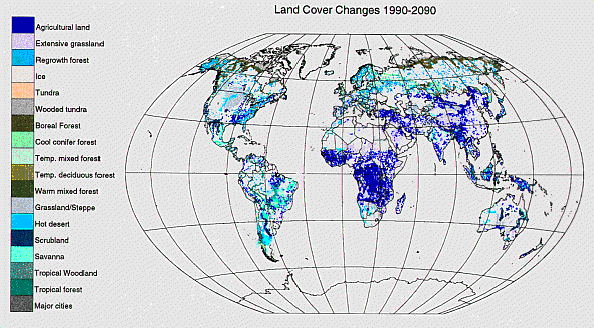
土地用途/覆被变化如在简介在世界范围内,至少在未来30年里,全球人口将以每10年约10亿的速度增长,粮食产量将需要以每年2%的速度持续增长。根据IMAGE模型(Alcamo 1994)和其他分析的预测,这将导致自然生态系统进一步向农业转变,特别是在非洲和亚洲(见图5),以及在现有耕作土地上集约化生产。这两个过程几乎总是会加速向大气中释放碳,因此这一来源的总体排放量至少会保持在当前水平,或者更有可能会增加。此外,随着更多的土地被转化为农业,自然生态系统中可作为碳汇的面积减少,从而降低了陆地生物圈的潜在碳汇强度。
图5所示。IMAGE 2.1模式对1990 - 2090年全球土地覆盖变化的预测彩色区域描绘了从一种封面类型转换为所示封面类型的区域。(出自R. Leemans,个人沟通).
生态系统结构的变化
生态系统组成和结构的变化是由管理实践(包括缺乏管理实践)和气候和大气组成的变化共同推动的。例如,目前在许多森林地区观察到的生物量增加在很大程度上反映了由于过去森林管理的变化而发生的连续变化。未来全球变化的影响将叠加在目前的趋势上。特别是,由于全球变化的影响日益重要,目前生物量增加的趋势在某些地区可能会逆转。在全球变化下,现有的植被组合,而不是作为完整的生物群落发生变化,可能会通过其某些组成部分的死亡率增加而发生变化,随后是新的组合的建立和生长。现有的植被向大气中释放碳,其死亡是一个快速的过程,而从大气中吸收碳的新植被组合的生长则较慢。因此,生态系统结构和组成变化的过程可能会导致在几十年到几百年的时间尺度上向大气中排放的碳增加,而不管新的理论平衡生物群落分布(假设未来气候稳定)最终存储的碳比目前分布多还是少。
生态系统生理学可能影响陆地生态系统长期碳平衡的具体生理因素包括:
土壤排放。据预测,随着气温的升高,土壤有机质的氧化程度将会增加。在高纬度地区,大陆地区在过去三十年里温度一直在上升,从这些地区的观测结果表明,阿拉斯加和西伯利亚的一些苔原生态系统已经从碳汇变成了碳源,或者大致处于平衡状态,这主要是由于土壤碳分解的增加。然而,在土壤排放作为一个新兴CO来源的潜在重要性之前,还需要做更多的工作,例如在几个生物群落中进行一组协调的变暖实验2可以确认。
有限公司2受精/ N沉积。最新生态系统级CO的综合2研究表明,CO的作用2对生态系统功能的影响,虽然仍然具有潜在的重要意义,但并不像之前想象的那么大。土地利用变化(退耕还林)、氮沉降和CO的交互作用2施肥导致了许多温带森林的强劲生长,有证据表明N沉降起了主要作用(例如,Auclair和Bedford 1995)。然而,氮沉降效应有一个明确的最大值,而且似乎在一些欧洲森林中超过了这个最大值(Schulze 1989)。
营养限制。在多个时间尺度上,碳循环与其他营养物质,特别是氮、磷和硫的循环密切而必然地联系在一起(Rastetter et al. 1997)。营养供应不足限制了许多系统的生态系统级碳吸收和储存,从而减弱了大气CO增加的影响2以及气候变化。在一些生物群落中,温度升高会导致N矿化增加,这本身会导致CO的增加2吸收,至少在短期内。
生理上的“饱和”。生态系统生理对大气中碳的净吸收是二氧化碳同化的平衡2通过光合作用和CO的释放2通过呼吸和分解。这些过程以不同的速度发生。碳同化对大气CO的增加几乎立即做出积极的反应2,而分解过程只是间接响应,通过温度、湿度和凋落物质量的变化,所有这些都包括长延迟成分。此外,这些过程还存在非线性。尽管碳同化随着大气CO的增加而增加2在美国,这种影响正在减弱。另一方面,呼吸作用是温度的指数增长函数。因此,随着全球变化的进行,CO的增长率2陆地生态系统的同化将会减缓,而呼吸和分解的速度将会加快。在短期内,这将对增长产生积极影响,从而对CO产生积极影响2但在更长的时间框架内(几个世纪),净效应将是陆地生物圈吸收CO的能力下降2.
总体趋势陆地碳预算中的各项不是独立运行的,也不是在同一时间尺度上运行的,这些特征使得很难从对短期尺度上一个预算项动态的了解推断出碳储量的长期趋势。净生物群系生产力的概念,NBP (Schulze和Heimann,在新闻),是整合多个过程在多个时间尺度上的影响的有用工具(图6).
图6所示。陆地生态系统碳的吸收和储存。
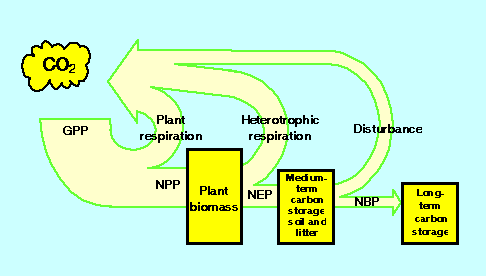
GPP,初级生产总值;净初级生产;NEP,净生态系统生产;NBP,净生物群系产量。
此外,预算中的条款在世界所有区域的作用并不相同。在一些地区,如撒哈拉以南非洲和亚洲大部分地区,土地利用变化可能占主导地位,这些地区将成为碳的净来源。对于其他地区,如北美和欧洲的部分地区,碳封存过程可能占主导地位,这些地区可能在十年或一个世纪的时间框架内保持或成为重要的碳汇。
考虑到很难估计未来(甚至现在)封存碳的过程(例如N沉积、CO2施肥)与释放碳(土壤有机质氧化、生态系统结构变化)相比,未来100年全球范围内陆地碳循环有许多可能的情景。然而,毫无疑问的是,至少在未来几十年里,更多的自然生态系统将转变为农业,同时排放碳,减少可以吸收大量碳的土地数量。因此,综合GCTE的总体结论是,在全球范围内,目前从大气中吸收碳的速度将难以维持。更有可能的是,陆地生物圈作为一个整体(包括为生产食物和纤维而改造或改造的土地)将成为一个净来源。这一预测对制定稳定大气中温室气体浓度的战略具有重大意义。
满足每年2%或更高的粮食需求增长的需要将给有管理的生产系统带来巨大压力。气候变化可能会进一步加剧这些系统的压力。极端天气事件,如一个连续干旱,或两个或两个以上的世界主要粮食产区同时干旱,将造成严重的粮食短缺。(美国中部持续的干旱。20世纪30年代的粮区造成了大规模的环境和经济破坏)。
解决粮食供应问题的方法之一是技术:发展改良的种植制度和/或作物品种。毫无疑问,改良品种,如含有内置杀虫剂的转基因作物和水分利用效率高的短季品种,将抵消部分增加的需求。然而,生物技术还没有成功地提高我们处理复杂的系统级问题的能力,例如干旱和盐度。几乎可以肯定,全球产量每年持续增长2%是可以实现的,但这将对土地利用和总体生态系统产生重大影响。beplay竞技气候变化使得生产额外食物和纤维的任务更加不确定。
资源的可得性将继续限制许多区域的农业发展。例如,由于农业、工业和城市对水的需求与为保护、清除废物和净化目的而维持河流流量的需求之间的直接竞争,已经成为一个主要问题的水资源供应可能会越来越受到限制。
在全球变化对陆地生产系统的影响以及对区域和全球粮食供应的影响方面,过去六年来的工作突出了下列主要问题:
-
•作物生产。世界不同地区的作物生产将受到非常不同的影响(最近的其他评估已经强调了这一点)。最近的估计表明,中高纬度地区产量增加,但低纬度地区产量减少,而低纬度地区的粮食需求将最大。
在理想的田间条件下,当前CO增加一倍,小麦产量不太可能增加10%以上2浓度;在平均管理条件下,5-7%的增长更为现实(Pinter et al. 1996)。主要的小麦模型正在迅速完善,但在使用任何单一模型进行空间外推时仍需谨慎表2).
表2。GCTE小麦网络的8个模型对小麦产量(1毫克= 1公吨)的最大和最小估计。模型使用常见的天气数据集和叶面积指数(LAI)发展的同一时间过程来模拟假设的小麦作物的生长。(来自Goudriaan et al. 1994)。
| 开花的日子 | 到期日 | 赖 (m2/ m2) |
地上总干质量(Mg/ha) | 谷物干质量 (毫克/公顷) |
|
| Crookston,明尼苏达州,美国(春小麦) | |||||
| 规定 | 183 | 216 | |||
| 最低 | 182 | 215 | 4.5 | 10.6 | 3.6 |
| 最大 | 187 | 216 | 4.5 | 16.1 | 8.8 |
| 莱利斯塔德,荷兰(冬小麦) | |||||
| 规定 | 166 | 207 | |||
| 最低 | 164 | 204 | 7.5 | 13.5 | 5.5 |
| 最大 | 166 | 207 | 7.5 | 26.4 | 12.1 |
-
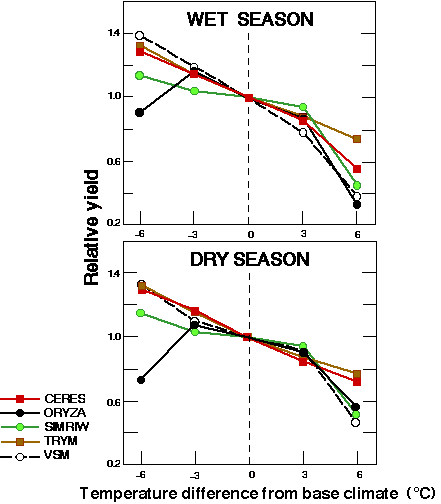
温度> 32oC由于小穗不育降低水稻产量。这种关系不受CO升高的影响2.主要的水稻模型在很大的潜在产量范围内一致,并建议每1个水稻产量减少5%左右oC上升温度> 32oC(见图7).
图7所示。由温度变化引起的水稻产量的相对变化,如五个生长模型所模拟的。(摘自Mitchell 1996)。
-
虫害、疾病和杂草目前造成作物产量的三分之一到一半的损失。beplay竞技气候变化可能导致热带和亚热带物种向温带地区扩散,并增加目前受高纬度地区热积累限制的物种的活动。
- •牧场和牧场。主要由于人口压力,作物/牧场边界将侵占发展中国家的牧场。牧场牲畜生产的变化主要是由于种植和蒸散量和降水的变化造成的土地面积减少。当前CO翻倍2在不同的牧场和牧场,根据温度、水分和养分限制,产量将提高0%至20%。对一个亚热带牧草的敏感性分析表明,CO使牧草生长增加了5%2通过减少年间NPP的变异性,可使牛的长期平均活重增加3%。
- •森林管理。CO升高的短期研究2在管理的森林中,在肥沃的条件下生长的幼树显示植物生物量产量的增加。然而,由于生态系统净呼吸的增加,从长期来看,这种增加将会减少。这种减少可能被CO的相互作用所大量补偿2大气氮沉降;净效果是不确定的。
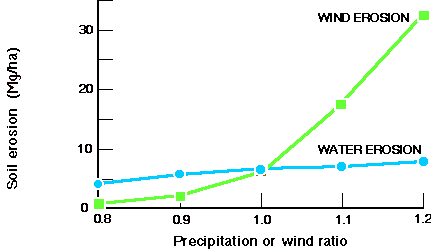
- •的土壤。在未来几十年里,控制土壤有机质(SOM)水平和侵蚀的主要方法仍将是土地管理,但近期气候变率变化导致的植被覆盖变化将是重大的,特别是在半干旱地区。平均温度的长期上升将加速SOM的氧化,特别是在它允许新的土地被种植的地方。土壤侵蚀与平均降水量呈线性增加,但与风速呈非线性增加,风蚀快速而显著增加的阈值约为5.5米/秒(见图8).
图8所示。利用EPIC模型估算的美国玉米带土壤侵蚀对气候变化的敏感性。beplay竞技每个点代表100个随机选择的地点的100年平均值。
生物多样性
最近对世界主要生物群落未来生物多样性趋势的分析(Sala和Chapin,在准备中)指出,未来几十年生物多样性丧失的主要原因是土地利用的变化,主要是栖息地丧失和景观破碎化。第二个最重要的因素是外来物种的入侵。虽然这里的趋势不太确定,但总的结论是,外来物种将是一个日益严重的问题,考虑到:(1)经济全球化,以及由此而来的人口和物质流动;(2)受干扰的生态系统对入侵的易感性。大气成分和气候的变化被认为是长期因素,随着时间的推移其相对重要性逐渐增加。然而,氮沉积的变化在较短的时间内对物种多样性产生重要影响,特别是在植物中,特别是在发达国家。
全球变化正在发生,在可预见的未来还将继续,并可能在许多方面加剧。这是一个正在出现的现实,将对政治进程、区域战略规划和资源管理人员的日常生活产生越来越大的影响。学会与全球变化共存,避免最坏的危险,并利用出现的机会,需要有创造性和创新的战略。这些必须建立在对陆地生态系统与全球变化相互作用的合理、科学的理解之上。
社会如何能够或应该利用现有的科学认识来应对全球变化?这种困难的一个例子是目前关于限制温室气体排放的“现在采取行动vs.以后采取行动(或不采取行动)”提案的辩论。一些全球进程有几十年甚至几百年的滞后时间。因此,现在不采取行动的后果可能要到下世纪中叶才能感受到,但当这些后果真的发生时,它们可能是严重的,非常难以应对。这种滞后效应的一个例子是陆地生物圈吸收碳作为大气CO的能力不断减弱2浓度和温度升高。现在不采取行动可能会导致一个巨大的,不可避免的,额外的CO2从现在开始一个世纪左右,通过增加土壤有机质的分解(在陆地生态系统中,地下储存的碳大约是地上储存的碳的两倍)来释放。
最重要的是,我们可能永远无法高度准确地预测陆地生态系统将如何与加速的环境变化相互作用。因此,认为生态系统可以像简单得多的人类设计的工业系统那样被“管理”的类比是误导性和危险的。就陆地生态系统与全球变化的相互作用而言,我们必须预料到意外(和不可预测),并保持尽可能多的应对选项。在农业生态系统中,恢复力和产量之间存在着不可避免的平衡:生产力最高的系统往往是最简单的,但它们对干扰和扰动的恢复力最差。需要高效率的系统来养活不断增长的人口;复杂、有弹性的系统需要能够应对未来的冲击和干扰,并继续提供我们所需的生态系统“商品和服务”。
欢迎对本文进行回复。如果被接受发表,您的回复将被超链接到文章。要提交评论,请遵循这个链接.阅读已接受的评论,请跟随这个链接.
致谢
这是国际地圈-生物圈计划(IGBP)的全球变化和陆地生态系统(GCTE)项目过去六年工作的详细综合概述。完整的综合报告将于1998年初由剑桥大学出版社出版,由30多位GCTE科学家撰写。本文作者根据IGBP秘书处于1997年12月在斯德哥尔摩分发的综合执行摘要编制了概述。
我们感谢德国国家IGBP秘书处和位于加利福尼亚州圣巴巴拉的国家生态分析和合成中心对1996年两次GCTE合成讲习班的慷慨财政支持。
阿尔卡莫,J.编辑。1994.图2.0:全球气候变化的综合建模。beplay竞技Kluwer学术出版社,多德雷赫特,荷兰。
Auclair, A.N.D和J.A. Bedford。1995.北方森林年净森林容量平衡的近期变化及其对全球碳平衡的影响。生态系统研究报告10、欧洲委员会,布鲁塞尔,卢森堡。
Goudriaan, J. S.C. van de Geijn和J.S.I. Ingram。1994.GCTE焦点3小麦模型和实验数据比较研讨会报告。英国牛津GCTE Focus 3办公室。
这个,M。1997.一个世纪前,阿伦尼乌斯和霍格博姆对当代全球碳循环的回顾。中记录26: 17-24。
IPCC不相上下。1995.beplay竞技气候变化1995 -气候变化科学。第一工作组对政府间气候变化专门委员会第二次评估报告的贡献。beplay竞技j·t·霍顿,l·g·梅拉·菲尔霍,b·a·卡兰德,n·哈里斯,a·卡滕伯格和k·马斯凯尔编辑。剑桥大学出版社,英国剑桥。
Korner, Ch。1995.为提高植物对高浓度CO的响应提供更好的实验依据2还有气候变暖。植物、细胞与环境18: 1101 - 1110。
库尔兹,w.a., M.J. Apps, S.J. Beukema和T. Lekstrum。1995.20世纪加拿大森林的碳预算。忒勒斯47个b: 170 - 177。
米切尔P.L.1996.5种水稻产量模型的比较,显示温度和二氧化碳浓度变化的影响。在编辑J.S.I.英格拉姆。GCTE水稻网络实验规划研讨会报告。GCTE工作文件
品特,P.J. Jr, b.a Kimball, R.L. Garcia, G.W. Wall, D.J. Hunsaker和R.L. LaMorte。1996.自由空间有限公司2富集:棉花和小麦作物的反应。215 - 249页在G.W.科赫和H. A.穆尼,编辑。二氧化碳和陆地生态系统。学术出版社,圣地亚哥,加利福尼亚,美国。
皮特尔卡,L.F.和植物迁移工作小组。1997.植物迁移与气候变化。beplay竞技美国科学家85: 464 - 473。
Rastetter, e.b., G.I. Ågren,还有G.R. Shaver。1997.n限制生态系统对CO增加的响应2:营养平衡,元素循环耦合模型。生态应用程序7: 444 - 460。
萨拉,o.e.,和F.S.查宾,三世,编辑。生物多样性丧失的未来情景。德国海德堡Springer-Verlag在出版社。
丢弃,科学博士1995.陆地生态系统和碳循环。全球变化生物学1: 77 - 91。
Schulze e d。1989.空气污染和云杉森林的减少(挪威云杉森林)。科学244: 776 - 783。
Schulze e d。和M.海曼。陆地生态系统碳水交换。在j·n·加洛韦和j·m·梅利洛,编辑。全球变化背景下的亚洲变化。IGBP丛书第3号,剑桥大学出版社,英国剑桥在出版社。.
沃克,b.h., W.L. Steffen, J. Canadell和J.S.I.英格拉姆,编辑。全球变化对自然和管理生态系统的影响:GCTE和相关研究的综合。IGBP丛书第4号,剑桥大学出版社,英国剑桥在新闻.
通讯地址:
布莱恩•沃克
CSIRO野生动物和生态
GCTE核心项目办公室
Bellenden路,冈加林,ACT 2600
电话:61-02-6242-1742
传真:61-02-241-2362
Brian.Walker@dwe.csiro.au
*本文版权于2000年1月1日由美国生态学会转让给韧性联盟。
![]()