

版权所有©2013作者(s)。由韧性联盟授权在此发布。
去pdf本文的版本
去pdf本文的版本
以下是引用这篇文章的既定格式:
加瓦佐夫,K. S., A. Peringer, A. Buttler, F. Gillet, T. Spiegelberger. 2013。气候变化情景下瑞士汝拉山区草地-林地牧草生产动态beplay竞技生态和社会 18(1): 38。
http://dx.doi.org/10.5751/ES-04974-180138
研究,一部分的特别功能山区可持续土地利用实践:全球变化下生态系统动态、社会经济影响和政策影响的综合分析
加瓦佐夫,K. S., A. Peringer, A. Buttler, F. Gillet, T. Spiegelberger. 2013。气候变化情景下瑞士汝拉山区草地-林地牧草生产动态beplay竞技生态和社会 18(1): 38。
http://dx.doi.org/10.5751/ES-04974-180138
气候变化情景下瑞士汝拉山区草地-林地牧草生产动态beplay竞技
1洛桑理工学院建筑学院,土木与环境工程,生态系统ECOS实验室,2瑞士联邦森林、雪和景观研究所,3.斯图加特大学景观规划研究所4法国弗朗什-康特大学(法国)5山地生态系统研究中心
摘要
瑞士汝拉山脉的林傍系统是亚高山植被带的传统饲料和木材来源,但它们对土地利用和气候变化的脆弱性使其未来的可持续性岌岌可危。beplay竞技我们结合实验和建模方法来评估气候变化对草地-林地景观的影响。beplay竞技采用沿林冠盖度梯度采样的方法,对不同经营强度的林分草地的抗性潜力进行了研究。这一梯度从与集约化农业相关的无木牧场到与粗放农业相关的密林牧场。将这些生态系统移植到温暖和干燥的条件下的中生态系统提供了实验证据,表明气候变化减少了非森林牧场的草本生物量生产,但对稀疏森林牧场没有影响,甚至刺激了茂密森林牧场的生产力。beplay竞技通过使用针对当前应用修改的森林牧场(WoodPaM)空间显式模型对这些结果进行建模,将结果外推到政府间气候变化专门委员会(ipcc)针对气候变化的两个区区化情景下的当地景观。beplay竞技这导致人们认为,在不久的将来(公元2000-2050年),Jura牧场-林地的牧草生产将在整个景观中受到不成比例的影响。在炎热、干燥的年份,只有通过广泛和适度的农业才能提供稳定的草料供应,这样就可以在放牧的牧场内形成绝缘的树木覆盖层。我们的结论是,这种结构景观多样性将赋予森林牧场在未来几十年面对气候变化时具有缓冲潜力。beplay竞技
关键词:地上生物量;干旱;群落交错区;草原;牧场;silvopastoral系统;亚高山带的;移植;林地
介绍
牧场林地的历史和重要性
草原林地是欧洲山区的一种传统的人工景观(Etienne 1996),覆盖了瑞士汝拉山脉的大部分。它们由开阔的草地、封闭的森林和半森林牧场的马赛克拼接而成,其中有分散或丛生的树木,其形状得益于长期的多功能土地利用历史,主要是放牧和林业(Buttler et al. 2009)。森林牧场是封闭森林和开放草地之间真正的过渡带,因此不仅仅是这两种植被类型之间的简单界面。在这种林带生态系统中,草原和林地在空间和时间上密切相关,这是相互抵消的生态过程之间的平衡的结果(Gillet et al. 2002, Gillet 2008)。森林入侵和牲畜放牧和广泛的牧场管理导致了景观的时空异质性,由扰动机制、微气候和地形定义,并为高生物多样性提供了有利条件(Gillet et al. 1999, Dufour et al. 2006)。目前,牧场-林地所提供的生态系统服务的数量和质量差别很大。经济上重要的因素,如饲料供应,通过集中管理的无树牧场比通过广泛的有自由放牧牲畜的森林牧场保持较高的水平。这对当地农民来说很重要,从社会经济的角度来看,他们是树木繁茂的牧场的主要使用者,因此,他们可能会故意将景观塑造成一个没有树木的景观。除了农业以外,在一些区域,林业活动产生的收入可能相当可观(Gillet和Gallandat 1996年)。这一景观对旅游经济的重要性也得到了承认,尽管它仍然难以衡量(Miéville-Ott和Barbezat 2005)。
在单一而多样的林带生态系统中,草场和林地的共存是不容易保护的,需要综合管理方案(Barbezat和Boquet 2008)。当前土地利用的集约化和扩大化都可能导致这种不稳定的联系的破裂,导致林地和草地的分离(Buttler et al. 2009)。尽管人为土地利用变化被认为是全球山地生态系统的一个主要威胁(Körner et al. 2006),但人们对其与未来气候变化的相互作用知之甚少。beplay竞技
beplay竞技欧洲山区的气候变化
据预测,全球变暖和相关的环境变化将在本世纪对山区生态系统产生强烈影响(IPCC 2007年)。20世纪在瑞士山区观测到的年平均气温上升1.5 K的趋势(Beniston等人,1997年)与极端夏季温度峰值增加和干旱延长的情况相一致(Schär等人,2004年,Beniston 2009年)。预计在整个本世纪,这些趋势将变得更加明显,北半球温带山脉将经历最强烈的温度上升,升温速率通常比20世纪记录的高2到3倍(范围为+2.8 K到+5.3 K) (Nogues-Bravo等人,2007年)。此外,在生长季预测的炎热天气和30%的降水减少将加剧中欧的干旱期(Beniston等,2007,CH2011)。这些将带来夏季土壤水分有效性、植物物候和生长季节长度的变化,最终将对生态系统的分布和功能产生影响(Parry 2000, FOEN/FSO 2011)。植物群落对气候变化的响应beplay竞技
偶发性极端气候事件,如夏季热浪,在景观尺度上具有强烈而独特的影响(Ciais等人,2005年,Reichstein等人,2007年,Teuling等人,2010年)。因此,对异质景观(如牧场-林地)中发生的初级生态过程的机制理解对于有效管理至关重要。目前,有大量的文献,包括各种实验方法,关于温暖和干燥气候对原始适应寒冷的植被的影响。沿着自然气候梯度的移植实验为检验关于物种和群落如何受未来气候变化影响的假设提供了一种强有力的方法。尽管使用该技术的已发表研究相对较少,但已经进行了一些成功的对比陆地环境的尝试:泥炭地(Wieder和Yavitt 1994, Breeuwer等人2010)、针叶林(Hobbie和Chapin 1998)、山地草甸(Bruelheide 2003)、亚高寒草地(Sebastia 2007)和高山植被(Scheepens等人2010)。然而,这些研究的结果勾勒出了植物群落对环境变化反应的特殊模式。关于植物地上生物量,最近的研究表明,随着气候变暖和变干燥,草本植物的年生物量产量会增加(Bruelheide 2003, Sebastia 2007)。其他人(Harte和Shaw 1995, Zhang和Welker 1996)则相反,他们没有观察到地上生物量的累积变化。仅干旱就已被发现对植物生物量产生负面影响(Johnson et al. 2011)。在植物群落组成的层面上,这些研究未能给出气候操纵导致的多样性变化的整体图景。 Either graminoids or forbs have been shown to successfully dominate after a climatic perturbation. Opportunistic plants may be granted competitive advantage through advanced phenological development (Dunne et al. 2003, Körner 2003, Inouye 2008), or through utilization of newly available nutrient resources (Bowman et al. 2006, Soudzilovskaia et al. 2007). Other plants may benefit from their inherent tolerance of specific environmental stress (Buchner and Neuner 2003, Brock and Galen 2005). Overall, more fertile early successional grassland communities have been shown to be more responsive to climate warming and drought, compared to late successional ones, typically found in areas of low intensity management (Grime et al. 2000). Drought resistance and recovery potential of plant communities have also been attributed to high species richness (van Ruijven and Berendse 2010, Mariotte et al. 2013), whereas grazing of highland pastures has been shown to both promote herbaceous richness and reduce evapotranspiration loss and thus water consumption (Körner et al. 2006). In comparison to grasslands, forests exhibit more conservative water use, and hence cope better with long-lasting heat and drought stress (Teuling et al. 2010).根据这些发现,并考虑到异质的景观结构和牧场-林地的高度生物多样性,我们怀疑这样一个森林和广泛放牧的山地草原群落的生产力可能对高温和干旱的影响是强有力的。
从实验地块升级到景观级别
Dunne等人(2004)的工作表明,将生态系统对气候变化的响应的实验结果从地块的规模外推断到景观的规模是一项具有挑战性但有益的任务,受此启发,我们在本研究中将实验与建模方法相结合。beplay竞技我们认为,这种观测、操作和建模技术的组合非常适合于评估生态系统的脆弱性或对环境变化的恢复力(Spiegelberger等,2012年)。为此,我们建立了一个沿着海拔梯度的移植实验,以获得开放草地、半森林牧场、我们将生物量生产的结果构建到森林-牧场生态系统的动态模拟模型中,以评估干旱事件在景观层面和气候变化预测的近时间框架(直到公元2050年)对牲畜饲料供应的影响。beplay竞技
假设
我们假设:(1)在气候变化下,森林草地的草本植物群落的地上生物量(AGB)生产比无树草地的更稳定。beplay竞技基于这一小区层面的假设,我们预测(2),在预计的热浪期间,森林牧场的景观尺度牧草生产将比集中管理的非森林牧场更加稳定。(3)在考虑未来气候变化影响的前提下,提出了森林牧场是一种比传统无树牧场更为稳健的土地利用形式的假说。beplay竞技方法
升温实验的研究区域和设计
位于瑞士Jura山脉的Combe des Amburnex是位于Jurasien Vaudois公园边界内的牧场-林地的特色亚高山地区。气候以海洋性为主,平均年降雨量约1750毫米,海拔1350米(米平均海拔高度)(包括超过450毫米的降雪),年平均温度4.5°C。从11月到5月,地面通常被雪覆盖。2009年8月,根据土地利用强度的不同,在海拔1350 m的Amburnex Combe des Amburnex沿线选择了三个面积相近的林地牧场。按管理强度的高低依次为:林木茂密的牧场、林木稀疏的牧场和无林木的牧场。这些牧场彼此之间的距离不超过1公里,每个牧场约2公顷,所有牧场的地貌、微地形和外貌都相同。在三种牧草类型中,随机选取15个样地代表各自区域的特色草本植被。为了移植土壤,地块被分为12个独立的20厘米× 20厘米和30厘米深的土壤芯,并组装回60厘米× 80厘米和30厘米高的PVC矩形盒子。从土芯中取出大块相交的树根(直径为> 1 cm),并用来自同一水平面的相邻土壤填充其余空隙。由此产生的45个中观群落被留在原地,直到植被季结束,从挖掘中恢复过来,然后在2009年10月被运送到它们的受体地点。在一个垂直样带上,总共建立了三个移植点:Combe des Amburnex (1350 m ASL, 46°54 ' N, 6°23 ' E)、Saint-George (1010 m ASL, 46°52 ' N, 6°26 ' E)和Arboretum d ' aubonne (ASL 570 m, 46°51 ' N, 6°37 ' E)。第一个移植点位于ASL 1350 m,作为气候不变的对照点。选择海拔1010 m的地点代表年气温上升+2 K和降水减少-20%的组合。海拔570 m的站点表现为年升温+4 K和降水减少-40%的组合。 These preliminary approximations were derived from interpolated data from nearby weather stations. At each site, 15 mesocosms, thus representing five replicates of each pasture type and all originating from the Combe des Amburnex, were transplanted following a completely randomized design. The boxes were dug down to surface level into previously prepared trenches, thus preventing lateral heat exchange with the atmosphere. Mesocosms with turfs from sparsely wooded pastures and densely wooded pastures were shaded using two types of UV-resistant nylon mesh, which reduced photosynthetic active radiation by 40% and 80%, respectively, thus simulating field-measured light conditions in the corresponding habitats during the 2009 growing season (未发表的数据).网状织物悬挂在距离地面50厘米的木框上,并没有拦截雨水的沉淀。他们只在一年中无雪的时候留在地里,以避免与积雪产生干扰。为了保持放牧对地块的压力,避免“废弃”和站立凋落物堆积的混杂效应,在植被生长季结束时,将地块修剪到接近地面的位置。
草本植物生物量于草坪移植后第一年的2010年7月底收获,并作为年度AGB产量的估算值。在每个地块,在35cm × 35cm的固定区域内,将植被砍至离土壤表面约1 cm,确定为物种水平,在70˚C下干燥48 h,并称重。
在整个实验过程中,通过每个试验点的一个自动气象站(Sensor Scope Sàrl, Switzerland)连续监测气候参数,该气象站每隔一分钟测量地面以上2米的空气温度和湿度,以及降水。决定自2O EC-TM传感器探头与Em50数据记录器(Decagon Devices, Inc., USA)耦合,每分钟记录表土层(0 ~ -3 cm)的土壤温度和体积含水量,并在一小时间隔内对数据进行平均。这里提供的数据是2010年生长季节的月份——4月、5月、6月、7月、8月和9月。
实验数据的统计分析
实验设计允许用四个因素解释移植后第一年草本植物生物量的年变化:两个分类因素——移植地点的初始牧草类型和海拔高度,以及两个连续的物理因素——土壤温度和土壤湿度。在生长季节(土壤解冻和收获日之间),每个地块的两个土壤变量都取平均值,从而综合了植物所经历的小气候条件。通过原始数据拟合ANCOVA模型,检验单因素的显著性及其双向作用。残差的正态性和同方差假设用诊断图进行了视觉验证。为了将在不同环境条件下获得的试验地数据外推到真实景观中连续的场地梯度,我们建立了一个线性回归模型,从树木覆盖率和温度变化程度这两个连续变量预测AGB的季节性产量。试验田遮阳与林木覆盖的关系如下:0%林木覆盖的非林木草地遮阳(冠下消光)为0%,50%林木覆盖的疏林草地遮阳为40%,100%林木覆盖的密林草地遮阳为80%。温度变化计算为试验年2010年当地植物生长季温度与对照地(海拔1350 m)温度之差。将保留在最优模型拟合中的这些因素的参数估计值纳入到WoodPAM的林地景观模型中。
所有统计分析均采用R 2.13.1版本进行。(R开发核心团队2012)。
森林-牧场生态系统空间显式模拟模型
Gillet et al.(2002)和Gillet(2008)建立了林间牧场动态模拟模型WoodPaM,用于研究瑞士汝拉山区林间牧场的演替动态。WoodPaM是一个空间显式的生态系统模型。因此,通过明确考虑从初级生产力到牲畜饲料消费的食物链,它能够模拟由于大型草食动物(牛)的选择性放牧而产生的半开放景观结构。在模拟过程中,牛的选择性觅食会对植被造成局部影响(即放牧、啃食、排泄和踩踏)。这些因素,再加上地貌和气候条件,促进或阻碍森林的发展,从而推动景观结构的发展。在WoodPaM中,牧场由25米宽的二次元网格单元表示(图1)。在每个单元中,植被由草本植物、灌木和树木子模型表示。草本层的演替是由放牧和粪便的强度以及树木覆盖所驱动的。因此,草本植被被分为以下四种植被类型:富营养化牧草、寡营养化牧草、休耕和林下植被,它们的牧区价值(PV)不同(表1)。基于新的回归模型(与Gillet[2008]比较)计算网格单元中牧草年产量“P”的初步估计,该研究将Gillet(2008)使用的来自Jura山脉和阿尔卑斯山的同一组植被调查的生产力经验数据与畜牧价值“PV”和植被生长期“vegdays”的长度联系起来,其中包括:
P =年牧草产量(t DM·ha)-1·一个-1)
PV =网格单元中草本层的平均牧值,以植被类型的相对覆盖度加权
植被生长期(天)
fpm =最大产量(20吨DM·公顷)-1·一个-1)
fpi = PV = 0时的“初始”产量(8.5 t DM·ha)-1·一个-1)
Ra =海拔对生长系数的影响(2.152·105)
rm = A = 0(0.05)时的最大生长系数
在模拟过程中,根据每个网格单元格中植被类型的当前牧值和过去50年模拟植被生长期的平均长度计算出年牧草产量的第一个估计值。通过这种方法,我们考虑到草地植物群落的生产力是随着气温上升的总体趋势(图2)而缓慢增长的,而不是在单个温暖年份自发增长的。在第二种情况下,干旱效应是可以预料的,我们单独讨论了这个问题。
与以前版本的WoodPaM相比,作为进一步的创新(见Gillet 2008, Peringer等人,2013),我们修改了饲草产量的第一个估计,方法是在炎热年份的植被生长期对平均高温的干旱影响应用乘数“D”。这一修正是根据AGB对移植响应的回归模型计算得出的,该模型沿气候变暖梯度(参见方法:统计分析而且结果:温度升高的影响).由于移植实验没有提供气候变冷的数据,所以在较冷的年份没有使用修饰剂。温度距平“ΔT”为1961-2010年植被生长期平均温度与植被生长期长期平均温度之差(实验发生在2010年,因此植物群落适应了最近的过去气候)。由于回归函数还表达了生产力对树木覆盖的依赖性(林下植被的牧草产量低于草地),因此将林下植被的牧值设为草地的值(表1)。根据公式1计算干旱和树木覆盖的综合影响:
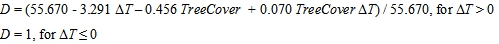 |
(1) |
模型修正假设:(1)近50年山地草地群落生产力随植被生长期的增加而缓慢增加(群落适应);然而,(2)在特别炎热的年份,它们的生产力持续受到干旱的影响。我们将当前气候(图2)作为干旱影响估计的基线,因为我们没有预料到新的(抗旱)基因型的发展,特别是因为草原是多年生物种的主宰,物种迁移缓慢。
因此,每个网格单元的模拟牧草产量是草本层的演替状态(植被类型的牧值)、近50年植被生长期的平均长度以及单个炎热年份的干旱影响的函数。网格细胞水平上的局部放牧影响取决于牛的选择性生境利用。反过来,网格单元对牛群放牧的吸引力取决于它们的饲料供应和当地的场地特征(即,地形的陡峭程度,岩石露头)。通过这种方法,建立了放牧影响与植被演替之间的闭环反馈,从而得到了模拟过程中草本层植被类型生产力的时空分布。
网格单元中消耗的牧草和牧草产量的比率用当地利用率(LUR)表示,并以每牲畜单位每天18公斤DM的牧草消耗为基础。在牧场水平上计算的相同比率称为全球利用率(GUR)。
有关模型结构和草本层和乔木层动态的实现以及选择性放牧规则的进一步详细描述,请参阅Gillet(2008)。Peringer等人(2013)给出了该模型描述之外的最新模型改进。
设计模拟
我们模拟了瑞士Bullet地区三个不同的森林牧场在观测和预测气候下的牧草生产。这些牧场位于瑞士汝拉山脉的同一景观中,与实验地点相同,沿山冠约40公里东北方向。这些牧场彼此相邻(图1);因此,它们有着相同的气候,但放牧强度不同,因此植被结构也不同。Les Planets Ouest(西经1200米,北纬46°83 ',东经6°55 ')是一个密集使用的公共地区,25.6公顷土地上有46个牲畜单位(LU),每年170天,饲养1.79公顷/公顷(另见Chételat等人,2013年)。这个牧场几乎没有树木,为了本研究的目的,被称为密集森林牧场(IWP)。Les Planets milu Est(西经1200米,北纬46°84′,东经6°55′)大部分是开阔的草原,但在遥远的西北部有一些树木稀疏的牧场。中度放牧密度为22公顷,占地14.1公顷,放牧天数为135天/年,放牧面积为1.56公顷/公顷,称为中度林地牧场(MWP)。Les Cluds Sud(西经1200米,北纬46°84′,东经6°56′)是一个马赛克牧场,有几片林地,广泛放牧23公顷,23.3公顷,每年153天,放牧0.99公顷。它被称为广泛的森林牧场(EWP)。 Although ongoing socioeconomic developments will likely lead to reduced stocking of wooded-pastures (Huber et al. 2013), for the purpose of this study, we simulated constant stocking densities.温度和降水的时间序列来源于对公元1950-2000年期间观测到的气候的分区和政府间气候变化专门委员会的两个气候变化情景beplay竞技特别报告排放情景(IPCC-SRES 2000)提供的公元2000 - 2100年的气候数据,该数据由英国诺里奇东安格利亚大学气候研究小组CRU和廷德尔气候变化研究中心(由D. Schmatz, wsll - switzerland,beplay竞技个人沟通).我们选择了温和情景B2和极端情景A1FI,假设现实的未来发展将在这两种情景之间的范围内。相应的年平均气温和植被生长期的年平均气温如图2所示。后者在模拟过程中驱动了干旱的影响,而过去50年的平均温度驱动了生产力的提高。我们从公元2000年的航拍照片初始化了模型,并应用了一个50年的旋转周期:公元1950-2000年。模拟从2000年开始持续了50年。
结果
温度升高对小区地上生物量产量的影响
移栽下坡使试验地的植物群落暴露在(预期的)环境气温升高和植被生长期降水减少的环境中(表2)。经过这种处理后,土壤温度普遍升高,而土壤湿度下降(表2)。在模拟树木覆盖的梯度上,每个地点的土壤温度对树冠遮荫水平(0%、40%和80%)的约1k几乎呈线性下降。与对照点相比,遮阳也能防止低海拔地区土壤水分蒸发。在密林牧场中,570 m ASL和1010 m ASL土壤水分含量分别保持在70%和96%。ANCOVA模型结果表明,在移栽梯度上,放牧类型对AGB产量有显著影响(P 2、25< 0.001),及其与移植部位海拔高度的相互作用(P 4、25< 0.001)。此外,土壤温度对其影响显著(P 1,25= 0.016)及土壤湿度(P 1,25= 0.008)。AGB沿土地利用强度梯度的顺序依次为非乔木草地>稀疏乔木草地>茂密乔木草地(图3)。这些差异在海拔1350 m的对照地点最为明显,但在较温暖的气候条件下减弱,在茂密乔木草地AGB产量增加,在非乔木草地减少,在稀疏乔木草地保持不变(见图3,以及牧草类型和海拔之间的统计显著交互作用)。仅移植海拔对各牧草类型间AGB的影响不一致。
利用生物量数据拟合线性回归模型,可以估计土地利用和气候参数对牧草生产力的影响。所得的模型(R 2= 0.85,P 3, 41< 0.001)在式2中给出:
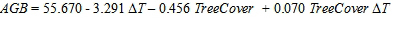 |
(2) |
这个方程表示草本生物量的产量,AGB,作为一个函数的树木覆盖率(TreeCover),空气温度异常(ΔT),以及他们之间的互动。所有的参数估计,包括模型的截距,都非常显著(P 1, 41< 0.001),因此被纳入WoodPaM模型,作为一个因子应用于草地-森林镶嵌的牧草产量的首次估计(方法:空间显式仿真模型).
景观层面温度异常对牧草生产的影响
在不同的放牧强度下,模拟的GUR时间序列遵循不同的轨迹(图4)。最明显的是较高的放牧密度和较高的利用率的细微影响,这导致每个牧场的曲线分离良好。模拟了粗放牧场(EWP: ~70%)、中等牧场(MWP: ~80%)和密集牧场(IWP: ~100%)的实际利用率。100%的利用率代表了经济意义上的最佳饲养密度,因为所有可用的饲料都被消耗掉了。如果利用率低于100%,则表明存在欠牧斑块,这是广泛放牧牧场的典型特征,并为灌木和森林的生长提供了生态位,从而形成特定的草地-林地景观马赛克。从2000年开始,利用率并没有下降,正如人们所预期的那样,随着气候变化,植被生长期延长,生产力提高(图2)。由于夏季炎热的年份增加,温度迅速上升,随后模拟干旱增加,弥补了这一影响。beplay竞技随着时间的推移,IWP和MWP的利用率开始波动,波动幅度越来越大,IWP的峰值超过了100%的阈值。相反,EWP的利用率始终保持在100%以下,波动也小得多。对于剧烈变暖情景A1FI (+8 K),这种影响甚至更加明显,特别是在模拟年2042-2047年预测的热浪中。
总体而言,就目前的饲养密度而言,对粗放牧场的模拟显示,牧草持续供应充足,而在密集的牧场,牧草短缺。由于该牧场的放牧密度本来就很低,因此在EWP中稳定的饲草供应(即图4中预测的GUR的低振幅)可能显得微不足道。尽管如此,在其森林景观斑块中,受刺激的AGB生产确实提供了对热浪的定量缓冲能力。通过对比IPCC-SRES气候变化情景B2模拟连续序列中的高温(2021年)和低温(2022年)两个模拟年份(图2),我们证明了景观异质性对生态系统过程恢复力的这种贡献。为了比较准相同的景观结构,我们选择了连续两年。beplay竞技在炎热的2021年,在没有树木的国际保护区,牧草产量下降,干旱导致GUR增加12.5%(表3,图5)。在MWP和EWP的草地上,可以发现类似的干旱对牧草产量的影响(图5);然而,eep(真正的森林-草地马赛克)的利用率仅在GUR上增加了6.7%(表3)。通过将给定年份的GUR值除以长期平均GUR值,对不太容易因牧草稀缺而超过GUR水平的广阔牧场的潜在偏差进行了额外的数学校正。两年间IWP和MWP的GUR值修正差异(分别为11.9和12.4)仍然高于EWP(9.5),这表明了模拟的森林牧场缓冲潜力(表3)。
讨论
气候变化对不同土地利用类型草本植物生产的beplay竞技显著影响
我们的实验研究是一项新颖的实验,可以直接比较气候变化对不同管理强度的邻近草地的影响,这些影响通过森林冠层盖度和草beplay竞技本物种组成的梯度表现出来。结果明确表明,在气候变暖的条件下,开放草地牧草生产力下降,而森林草地牧草生产力稳定甚至提高。这种模式在仅仅一年的实验处理后就变得明显,这强调了随机热浪对牧场-林地生态系统功能的重要性。由于移植处理的结果,露天牧场的干旱效应比两种木牧类型更强,这主要是由于土壤水分有效性下降。尽管我们对中尺度群落遮荫的简单设计当然没有包括真正树冠的所有小气候影响(即环境湿度、风拦截、树冠渗滤液、凋落物沉积或地下资源竞争),但我们相信,由于适当使用对照地块和整体较短的处理时间跨度,这防止了混杂的遗留效应积累,我们在草本植物生产方面的结果是可靠的。导致AGB下降的最显著因素之一是植物生长季节土壤水分有效性的下降。土壤温度随气候变暖呈线性增加,而非林地草地土壤湿度主要受影响,与之平行的是立木植物AGB的下降。此外,由于干旱,大部分AGB在生长高峰期过后不久就已经衰老(归一化植被指数测量,未发表的数据).这一现象的产生是由于与荫蔽地相比,开放林冠地的蒸散作用增强了,在荫蔽地,网格覆盖充分发挥了“森林般的”作用,在炎热的夏季限制了与大气的能量交换(Teuling et al. 2010)。
另一种解释可能是集约化农业(Körner et al. 2006),加上高施肥水平和受阻碍的连续发展(Grime et al. 2000),使集约化管理的开放牧场更容易受到气候变化的影响。beplay竞技人们可以推测,这些由快速生长的物种组成的草地群落对环境变化的反应更灵敏,它们能获得高的养分。然而,这种情况在我们的实验设计中没有得到明确的测试(没有施肥操作),仍然只是假设。此外,我们认为没有树木的牧场需要密集管理,但相对于有树木的牧场而言,这种管理更为严格,因为它们通常是亚高山牧场,而不是农田。
气候变化下牧草供给的景观格局beplay竞技
我们将实验结果纳入基于过程的森林牧场模拟模型的方法比跨空间尺度(从地块到景观水平)的简单外推更有价值。由于草地形态的生物量生产代表了WoodPaM模拟的基础,气候变化对初级生产的影响在机械上决定了牛的牧草可用性,而这反过来又通过选择性放牧行为塑造了景观结构(科勒等,2006年)。beplay竞技尽管我们的实验设计存在局限性,没有考虑到降水和温度之间看似合理的相互作用,但我们并不期望在我们的解释中存在定性偏差。我们的信心来自于当地观测到的和预测的这两个气候因素之间的强负相关;因此,我们在预测AGB产量的模型中只考虑了温度。尽管我们的移植梯度选择是基于当前的天气观测和气候预测,但我们无法解释生态系统对未来植物生长季节的反应,包括温暖和潮湿,或寒冷和干燥。其他关于气候变化对森林牧场影响的研究主要依赖于beplay竞技树木物种的气候敏感性(Peringer等人,2013年),而我们的研究关注的是草地的功能。模拟的设计涵盖了从公元2000年到2050年的更短的时间框架,这段时间足够检测到气候压力,同时假设没有植物物种适应(适应干旱的生态类型的发展),或通过移民造成的重大社区变化。尽管这一假设在过去几十年的模拟中可能显得不现实,但我们考虑了三个原因:(1)在模型中没有简单的方法来实现草地物种的进化和扩散过程;(2)由于森林对草本植物扩散的阻碍,气候变化对该地区低地物种迁移的影响可能非常缓慢beplay竞技;(3)由于山地牧草历史悠久(生态连续性),既有植物群体的表型可塑性和遗传多态性可能允许对干旱胁迫有一定的适应能力。此外,这段时间太短,无法对树木层和景观结构产生任何实质性的变化(关于气候变化的长期动态,请参见Peringer等人[2013])。beplay竞技然而,人们应该记住,从长远来看,气候变化对草地生产力的影响可能会由于牛的适应性放牧行为而对景观结构产生反馈(Smit et al. 2007, beplay竞技Vandenberghe et al. 2007, Vandenberghe et al. 2009)。在炎热的年份,露天牧场的牧草资源枯竭会促使牛群在森林下层植被中放牧,那里的干旱影响较小,牧草仍然可用。因此,森林的再生将受到威胁,景观结构动态可能会转向更开放的景观。这样的发展将抵消预期的森林侵蚀和冠层增厚(Peringer等人,2013年),从而在遥远的未来提供稳定的饲料供应。
我们的模拟结果表明,在粗放放牧系统中,不仅是普遍较低的放养密度,而且由此产生的草地-森林镶嵌有助于提供健壮的饲料。疏林草地AGB产量稳定,密林草地AGB产量增加,与非林草地AGB产量下降形成鲜明对比。因此,在生产力方面,无木牧草的明显优势随着干旱而减少,而有木牧草成为同样重要的饲料来源。鉴于稀疏和浓密的森林牧场覆盖在广泛放养的牧场上的面积相对较大,这种以前不高产的斑块显示出明显的潜力,可以弥补开放草地生产力的下降。
鉴于马赛克的出现是一个持续数世纪的过程(Gillet 2008, Peringer et al. 2013),系统的这种韧性是一种宝贵的商品,因为它会迅速被破坏,但会缓慢地重新建立。Chételat等人(2013)表明,如此巨大的景观变化可能是由更高的木材和食物需求(例如,在战争时期)和自然事件(风暴、干旱、树皮甲虫爆发)造成的。关于干旱期间树冠对林下小气候的有益作用,农林业系统也得出了类似的结论(Powell和Bork, 2006年)。在整个景观中,空间变化本身会刺激生态系统的恢复力和抗旱性。Godfree等人(2011)表明,通过异质环境的保护作用,干旱等自然发生的极端气候事件可以得到缓解。Foley等人(2005)也认同这一概念,他们主张通过单一的生态系统来维护多样化的生态系统服务组合,这样可持续的土地利用战略就可以同时满足短期和长期的需求。我们表明,在亚高山森林牧场的情况下,这可以通过马赛克景观的保存来实现。然而,人们承认,关于文化景观(即森林牧场)的未来的决定来自利益相关者而不是科学家,这种稳定性既需要维护历史上确立的农业实践(Küster 2004),也需要适应性管理,包括政策的改变(Huber et al. 2013)。无论是强化还是放弃,都将危及生态系统的稳定,并破坏其主要生态系统服务的提供。
结论
综上所述,瑞士汝拉山区草地-林地景观中的不同草地类型在草本牧草供给方面对气候变化表现出严格的非均匀响应。beplay竞技面对气候变暖,景观的马赛克斑块将是一项宝贵的资产,其固有的多样性可能是可持续土地利用管理的关键。提出了经验和模型证据,表明森林牧场可以在未来几十年缓冲气候变化的影响,以强有力的方式为牲畜提供饲料。beplay竞技我们认为,景观的这一属性应该被计入,以避免未来的景观隔离和相关的经济和文化影响。
致谢
这项工作得到了CCES(瑞士ETH领域环境和可持续性能力中心)作为mounland项目的一部分的支持,以及教育和研究国务秘书处(SER C07.0112)在成本行动FP0603框架下的支持。我们感谢国家奥波恩植物园、圣乔治公社(VD)、侏罗纪公园、Chamblard基金会对后勤和基础设施的支持。特别感谢西尔瓦娜·西霍夫博士,她为我们的手稿提供了建设性的意见,并感谢ECOS实验室的许多人在实地工作中给予了宝贵的帮助。
文献引用
Barbezat, V.和J. F. Boquet. 2008。建议intégrée des paysages sylvoo -pastoraux de l'Arc jurassien-Manuel(手册)。TransJurassienne会议。贝桑松La Chaux-de-Fonds上。
Beniston m . 2009。瑞士气候变量概率分布函数尾部的十年尺度变化。国际气候学杂志29:1362 - 1368。http://dx.doi.org/10.1002/joc.1793
贝尼斯顿,M. H. F.迪亚兹,R. S.布拉德利,1997。高海拔地区的气候变化:概述。气候变化36:233 - 251。http://dx.doi.org/10.1023/A:1005380714349
Beniston, M. D. B. Stephenson, O. B. Christensen, C. A. T. Ferro, C. Frei, S. Goyette, K. Halsnaes, T. Holt, K. Jylha, B. Koffi, J. Palutikof, R. Scholl, T. Semmler,和K. Woth. 2007。欧洲气候未来极端事件:区域气候模式预测的探索。气候变化81:71 - 95。http://dx.doi.org/10.1007/s10584-006-9226-z
鲍曼,W. D., J. R.高德纳,K.霍兰德和M.维德曼。2006。高山植被氮临界负荷与陆地生态系统响应:我们到了吗?生态应用程序16:1183 - 1193。http://dx.doi.org/http: / / dx.doi.org/10.1890/1051 - 0761 (2006) 016 (1183: NCLFAV) 2.0.CO; 2
B. J. M. Robroek和F. Berendse. 2010。全球变化的现场模拟:向南迁移北部沼泽中观生态系统。生态系统13:712 - 726。http://dx.doi.org/10.1007/s10021-010-9349-y
布洛克,M. T.和C.盖伦,2005。高山蒲公英的耐旱性,蒲公英ceratophorum(菊科),它的异域同族t . officinale自然和实验条件下的种间杂交。美国植物学杂志》92:1311 - 1321。http://dx.doi.org/10.3732/ajb.92.8.1311
Bruelheide, h . 2003。山地草甸的迁移以模拟气候变化的潜在影响。beplay竞技应用植物科学6:23-34。http://dx.doi.org/10.1111/j.1654-109X.2003.tb00561.x
Buchner, O.和G. Neuner. 2003。不同海拔高寒植物物种耐热性的变异。北极、南极和高山研究35:411 - 420。http://dx.doi.org/10.1657/1523 - 0430 (2003) 035 (0411: VOHTIA) 2.0.CO; 2
巴特勒,A. F.科勒和F.吉列,2009。瑞士山地森林牧场:模式和过程。377 - 396页在A. Rigueiro-Rodrigues, J. McAdam和M. R. Mosquera-Losada,编辑。欧洲农林业的现状和未来展望.施普林格,荷兰。http://dx.doi.org/10.1007/978-1-4020-8272-6_19
CH2011。2011.瑞士气候变化beplay竞技情景CH2011.C2SM,瑞士气象局,ETH, NCCR气候和OcCC,瑞士苏黎世。
陈特拉特,J. M.卡尔伯马滕,K. S. M.兰纳斯,T. Spiegelberger, J. B. Wettstein, F. Gillet, A. Peringer和A. Buttler. 2013。瑞士汝拉山区两个森林牧场土地利用和植被变化的背景分析。生态和社会18(1): 39。http://dx.doi.org/10.5751/ES-05287-180139
希艾斯,P, M. Reichstein, N. Viovy, A. Granier, J. Ogee, V. Allard, M. Aubinet, N. Buchmann, C. Bernhofer, A. Carrara, F. Chevallier, N. De Noblet, A. d Friend, P. Friedlingstein, T. Grunwald, B. Heinesch, P. Keronen, A. Knohl, G. Krinner, D. Loustau, G. Manca, G. Matteucci, F. Miglietta, J. M. Ourcival, D. Papale, K. Pilegaard, S. Rambal, G. Seufert, J. F. Soussana, M. J. Sanz, E. D. Schulze, T. Vesala, R. Valentini. 2005。2003年的高温和干旱导致全欧洲初级生产力下降。自然437:529 - 533。http://dx.doi.org/10.1038/nature03972
杜福尔,A., F. Gadallah, H. H. Wagner, A. Guisan和A. Buttler. 2006。山地景观植物物种丰富度与环境异质性:变异与空间构型的影响。描述生态学29:573 - 584。http://dx.doi.org/10.1111/j.0906-7590.2006.04605.x
邓恩,J. A.哈特,K. J.泰勒,2003。亚高山草甸开花物候学对气候变化的响应:综合实验和梯度方法。beplay竞技生态专著73:69 - 86。http://dx.doi.org/10.1890/0012 - 9615 (2003) 073 (0069: SMFPRT) 2.0.CO; 2
邓恩,j.a., S. R. Saleska, M. L.费舍尔和J.哈特,2004。生态气候变化研究中实验方法与梯度方法的结合。beplay竞技生态85:904 - 916。http://dx.doi.org/10.1890/03-8003
艾蒂安,M,编辑器。1996.西欧的银腭系统.INRA Editions,巴黎,法国。
联邦环境和联邦统计局(FOEN/FSO)。2011.环境2011年瑞士.伯尔尼和瑞士Neuchâtel。
Foley, J. A, R. DeFries, G. P. Asner, C. Barford, G. Bonan, S. R. Carpenter, F. S. Chapin, M. T. Coe, G. C. Daily, H. K. Gibbs, J. H. Helkowski, T. Holloway, E. A. Howard, C. J. Kucharik, C. Monfreda, J. A. Patz, I. C. Prentice, N. Ramankutty, P. K. Snyder, 2005。土地使用的全球后果。科学309:570 - 574。http://dx.doi.org/10.1126/science.1111772
吉莱,2008年成立。非均质草地-林地景观植被动态模拟。生态模型217:1-18。http://dx.doi.org/10.1016/j.ecolmodel.2008.05.013
吉列,F。o。贝松和j。m。Gobat》2002。森林草地植被动态的隔室模型。生态模型147:267 - 290。http://dx.doi.org/10.1016/s0304 - 3800 (01) 00427 - 6
吉列,F.和J. D.加兰达。1996。侏罗山脉树木繁茂的牧场。页面37-53在m . Etienne编辑器。西欧的银腭系统.INRA Editions,巴黎,法国。
吉列,F., B.穆里榭,A.巴特勒,j . d。Gallandat和人类。Gobat》1999。亚高山草甸乔木盖度对草本群落多样性的影响应用植物科学2:47-54。http://dx.doi.org/10.2307/1478880
Godfree, R, B. Lepschi, A.居里,T. Bolger, B. Robertson, D. Marshall, M. Carnegie, 2011。多尺度地形异质性增强了优势草地物种对极端干旱和气候变化的恢复力和抵抗力。beplay竞技全球变化生物学17:943 - 958。http://dx.doi.org/10.1111/j.1365-2486.2010.02292.x
格里姆,J. P.布朗,K.汤普森,G. J.马斯特斯,S. H.希利尔,I. P.克拉克,A. P.阿斯丘,D.考克,J. P.基尔蒂。2000。两个对比鲜明的石灰岩草地对模拟气候变化的响应。beplay竞技科学289:762 - 765。http://dx.doi.org/10.1126/science.289.5480.762
哈特,J.和R.肖,1995。山地植被群落中优势的转移——气候变暖实验的结果。科学267:876 - 880。http://dx.doi.org/10.1126/science.267.5199.876
霍布斯,S. E.和F. S.查宾三世。1998.北极冻土带林木生长极限试验研究。《生态学杂志》86:449 - 461。http://dx.doi.org/10.1046/j.1365-2745.1998.00278.x
Huber, R, S. Briner, A. Peringer, S. Lauber, R. Seidl, A. Widmer, F. Gillet, A. Buttler, Q. Beo Le,和C. Hirschi. 2013。草地-林地环境服务付费实施中的社会-生态反馈效应建模。生态和社会18卷。在新闻.
井上,d.w. 2008。气候变化对山地野花物候beplay竞技、霜冻危害和花丰度的影响。生态89:353 - 362。http://dx.doi.org/10.1890/06-2128.1
政府间气候变化专门委员会。beplay竞技2000.特别报告排放场景.剑桥大学出版社,瑞士日内瓦。
政府间气候变化专门委员会。beplay竞技2007.beplay竞技2007年气候变化:综合报告.第一、第二和第三工作组对政府间气候变化专门委员会第四次评估报告的贡献。beplay竞技瑞士日内瓦。
Johnson, J. Vachon, A. J. Britton, R. C. Helliwell. 2011。干旱通过对禾本科植物和蛙类植物的不同影响,改变了高山雪床生态系统的碳通量。新植物学家190:740 - 749。http://dx.doi.org/10.1111/j.1469-8137.2010.03613.x
科勒,F.吉列,S.雷斯特,H. H.瓦格纳,F.加达拉,J. M.戈巴特和A.巴特。2006。山地林木牧场牛生境利用的空间和季节格局景观生态学21:281 - 295。http://dx.doi.org/10.1007/s10980-005-0144-7
Korner, c . 2003。高山植物生命:高山生态系统的功能植物生态学.施普林格,柏林,海德堡,纽约。
Körner, C., G. Nakhutsrishvili,和E. Spehn. 2006。高海拔土地利用、生物多样性和生态系统功能。页面3-21在E. Spehn, M. Liberman和C. Körner,编辑。土地利用变化与山地生物多样性.CRC出版社,博卡拉顿,美国佛罗里达州。http://dx.doi.org/10.1201/9781420002874.ch1
工业,h . 2004。文化景观。页1 - 11在M.迪特里奇和J.斯特拉滕,编辑。文化景观与土地利用.施普林格,荷兰。
Mariotte, P., C. Vandenberghe, F. Hagedorn和A. Buttler. 2013。从属植物种类增强草地生态系统群落抗旱能力。《生态学杂志》 http://www.journalofecology.org/view/0/index.html
Miéville-Ott, V.和V. Barbezat. 2005。感知du pâturage boisé: résultats d'un sondage effectué au公社de La Sagne NE。德国日报:1 - 12。http://dx.doi.org/10.3188/szf.2005.0001
Nogués-Bravo, D. M. B. Araújo, M. P. Errea和J. P. Martinez-Rica。2007.21世纪全球山地系统对气候变暖的影响。全球环境变化-人与政策层面17:420 - 428。http://dx.doi.org/10.1016/j.gloenvcha.2006.11.007
帕里,2000年。欧洲气候变化的潜在影响和适应评估:欧洲ACACIA项目beplay竞技.英国诺里奇东安格利亚大学杰克逊环境研究所。
Peringer, S. Siehoff, J. Ch ? telat, T. Spiegelberger, A. Buttler, F. Gillet. 2013。气候变化下瑞士汝拉山区草地林地的过去与未来景观动态beplay竞技生态和社会18卷。在新闻.
鲍威尔,G. W.和E. W.博克,2006。白杨冠层去除和根沟对林下植被的影响。森林生态与经营230:79 - 90。http://dx.doi.org/10.1016/j.foreco.2006.04.021
R开发核心团队,2012。2.14.1 R版本。
Reichstein, M, P. Ciais, D. Papale, R. Valentini, S. Running, N. Viovy, W. Cramer, A. Granier, J. OgÉE, V. Allard, M. Aubinet, C. Bernhofer, N. Buchmann, A. Carrara, T. GrÜNwald, M. Heimann, B. Heinesch, A. Knohl, W. Kutsch, D. Loustau, G. Manca, G. Matteucci, F. Miglietta, J. M. Ourcival, K. Pilegaard, J. Pumpanen, S. Rambal, S. Schaphoff, G. Seufert, J. F. Soussana, M. J. Sanz, T. Vesala, M. Zhao. 2007。2003年欧洲夏季气候异常期间生态系统生产力和呼吸的减少:联合通量塔、遥感和建模分析。全球变化生物学13:634 - 651。http://dx.doi.org/10.1111/j.1365-2486.2006.01224.x
Schär, C. P. L. Vidale, D. Lûthi, C. Frei, C. Haberli, M. A. Liniger, C. Appenzeller. 2004。欧洲夏季热浪中不断增加的温度变化的作用。自然427:332 - 336。http://dx.doi.org/10.1038/nature02300
J. F.谢彭,E. S.弗雷,J.斯托克林。2010。广泛分布的高山植物移栽到不同海拔后比叶面积的基因型和环境变异。环境科学164:141 - 150。http://dx.doi.org/10.1007/s00442-010-1650-0
Sebastia M.-T。2007.植物行会驱动亚高寒草地的生物量对全球变暖和水资源有效性的响应。应用生态学杂志44:158 - 167。http://dx.doi.org/10.1111/j.1365-2664.2006.01232.x
Smit, C. Vandenberghe, J. den Ouden和H. Mûeller-Schäerer。2007.护理植物、树苗和放牧压力:沿生物环境梯度的促进作用变化。环境科学152:265 - 273。http://dx.doi.org/10.1007/s00442-006-0650-6
Soudzilovskaia, n.a, V. G. Onipchenko, J. H. C. Cornelissen和R. Aerts. 2007。施肥和灌溉对高山冻土带“叶生”的影响。植被科学杂志18:755 - 766。http://dx.doi.org/10.1111/j.1654-1103.2007.tb02591.x
Spiegelberger, T., F. Gillet, B. Amiaud, A. Thébault, P. Mariotte,和A. Buttler. 2012。植物群落生态学家如何考虑观察、实验和理论建模方法的互补性?植物生态学与进化145:4-12。http://dx.doi.org/http://dx.doi.org/10.5091/plecevo.2012.699
Teuling, a.j., s.i. Seneviratne, R. Stockli, M. Reichstein, E. Moors, P. Ciais, S. Luyssaert, B. van den Hurk, C. Ammann, C. Bernhofer, E. Dellwik, D. Gianelle, B. Gielen, T. Grunwald, K. Klumpp, L. Montagnani, C. Moureaux, M. Sottocornola, G. Wohlfahrt. 2010。欧洲森林和草原对热浪能量交换的对比响应。自然地球科学3:722 - 727。http://dx.doi.org/10.1038/ngeo950
范鲁吉文,J.和F. Berendse. 2010。干旱后,多样性促进了群落的恢复,但对抗旱性没有影响。《生态学杂志》98:81 - 86。http://dx.doi.org/10.1111/j.1365-2745.2009.01603.x
范登伯格(c.f .) Freléchoux;摩拉维,F. Gadallah和A. Buttler, 2007。牛采食对山地林木牧场树苗生长的短期影响。植物生态学188:253 - 264。http://dx.doi.org/10.1007/s11258-006-9160-1
Vandenberghe, C. Smit, M. Pohl, A. Buttler和F. Frelechoux. 2009。护理灌木的促进作用强度是否取决于树苗的抗放牧能力?基础与应用生态学10:427 - 436。http://dx.doi.org/http://dx.doi.org/10.1016/j.baae.2008.08.009
维德,R. K.和J. B.亚维特,1994。泥炭地与全球气候变化:来自位于纬度梯度的站点的比较研究的见解。湿地14:229 - 238。http://dx.doi.org/10.1007/BF03160660
张永强,张永强,韦克尔。1996。西藏高寒苔原对模拟气候变化的响应:地上生物量和群落响应。北极和高山研究28:203 - 209。http://dx.doi.org/10.2307/1551761
