

版权所有©2014作者(s)。由韧性联盟授权在此发布。
去pdf本文的版本
去pdf本文的版本
以下是引用这篇文章的既定格式:
Timmermann, K., G. E. Dinesen, S. Markager, L. Ravn-Jonsen, M. Bassompierre, E. Roth, J. G. Støttrup。2014.丹麦利姆峡湾(Limfjord)温带河口不同营养状况下贻贝渔业管理的生物经济模型的开发和使用。生态和社会 19(1): 14。
http://dx.doi.org/10.5751/ES-06041-190114
研究
Timmermann, K., G. E. Dinesen, S. Markager, L. Ravn-Jonsen, M. Bassompierre, E. Roth, J. G. Støttrup。2014.丹麦利姆峡湾(Limfjord)温带河口不同营养状况下贻贝渔业管理的生物经济模型的开发和使用。生态和社会 19(1): 14。
http://dx.doi.org/10.5751/ES-06041-190114
丹麦利姆峡湾(Limfjord)温带河口不同营养状况下贻贝渔业生物经济管理模型的开发与应用
1丹麦罗斯基勒奥尔胡斯大学生物科学系,2丹麦技术大学国家水产科学研究所海岸生态科,丹麦,夏洛特伦3.南丹麦大学环境与商业经济系,丹麦,埃斯布厄格,4诺和诺德公司,卡伦堡,丹麦
摘要
世界各地的沿海生态系统正面临着人为造成的营养输入、渔业活动、海水养殖、建筑工作和气候变化的压力。beplay竞技因此,综合管理手段必须结合社会经济问题处理其中一个或多个问题,以确保资源的可持续利用。在丹麦的一个温带富营养化河口利姆峡湾(Limfjord),有必要减少营养负荷,以履行诸如《水框架指令》(WFD)等欧盟法规。这些负荷减少的预期结果是改善水质,但也减少了大量滤食性蓝贻贝的产量,贝壳类.预计这将对利姆峡湾目前价值百万欧元的贻贝渔业产生重大的经济影响。我们开发了一个生物经济模型,可用于探索减少贻贝渔业目前的负荷的后果,以及潜在的管理选项,以获得经济和生态可持续的贻贝渔业。模型模拟清楚地表明,在达到WFD目标所需的营养负荷减少后,贻贝产量大幅下降。按照目前的做法,利姆峡湾的贻贝渔业在未来富营养化程度降低的河口将不会有利可图。然而,模型模拟也显示,贻贝渔业在实施世界捕捞日后,可以通过减少总捕捞配额、减少渔船数量和提高每艘渔船的捕捞配额来盈利。
关键词:决策支持系统;ecological-socioeconomic模型;富营养化;贻贝生产;营养负荷;预测模型;系统的管理;水框架指令
介绍
沿海地区是地球上人口最密集的地区,人类活动以各种方式给沿海海洋生态系统的完整性带来压力,例如营养物质的装载、密集的捕鱼、建筑工作和气候变化。beplay竞技管理这些相互作用的压力需要基于科学的方法,这些方法可以帮助实现资源的可持续利用,同时保护自然(诺贝尔奖2011年)。已经建立的管理系统通常在部门层面上控制这些压力,尽管已经尝试将多个生态系统组成部分包括在内,如实施渔业生态系统方法框架的指导方针(Garcia和Cochrane, 2005年)。这种方法鼓励了生物经济模型的发展,以探索不同物种的管理选项的生态和经济后果,如贝类(Imeson和Van den Bergh 2004)、剑鱼(Tserpes等人2009)和虾(Huang和Smith 2011)。这些模型构成了决策支持系统(DSS)的基础,DSS通过组织数据和以用户友好的方式提供模型和场景来帮助管理者(Geertman和Stillwell 2009, Nobre 2011)。一个实际实施的例子是为荷兰瓦登海开发的WadBOS系统,以获得系统的整体概述和改进管理(Engelen et al. 2003)。利用这一工具,从经济和生态后果方面探索了两种人类活动的新政策解决方案,贝类渔业和休闲划船,其中包括不同的利益攸关方(信息可在以下网站获得)http://www.riks.nl/Resources/papers/ManualWadBOSInternational.pdf).系统方法框架(SAF)是一种以整体方式解决政策问题的方法,由欧盟SPICOSA项目为2007年至2011年的沿海系统制定(Hopkins et al. 2011)。发展支助系统是武装部队的重要组成部分。SAF包括:(1)确定关键政策问题和利益相关者,(2)定义系统边界,以及(3)开发对这些问题至关重要的概念模型(Tett et al. 2011)。SAF建议管理策略应该由包括科学家、经理和利益相关者在内的团队来制定。它结合了基于生态、社会和经济过程现有数据的多学科科学分析(Hopkins et al. 2011)。注重用户友好的生物经济模型的发展促进了与利益相关者之间的沟通,是SAF在复杂系统中的实际应用的一个重要因素。这里提出的模型形成了丹麦北部一个温带富营养化河口利姆峡湾(Limfjord)的决策支持系统的核心。
利姆峡湾是复杂海岸系统的一个很好的例子,它受到多种人类活动的长期压力,导致了生态系统的重大变化。这些问题包括海水清晰度和水下植被的丧失(Krause-Jensen et al. 2012)、普遍的缺氧、底栖鱼类数量的锐减以及蓝贻贝生物量的同时增加,贝壳类Linnaeus, 1758 (Christiansen et al. 2006)认为这是浮游植物生物量高的结果。较高营养水平的变化导致了利姆峡湾(Limfjord)河口的开发同时发生变化,在那里以前繁荣的底栖鳍鱼渔业已被拖网捕捞蓝贻贝所取代(Dolmer和Frandsen, 2002年)。此外,有记录显示,20世纪90年代初发生了一种制度的转变,从底栖鱼类的主导地位转变为中上层鱼类、小型鱼类和无脊椎动物的主导地位(Tomczak等,2013年)。近年来,贻贝生物量和渔获量随着营养物负荷和浮游植物产量的下降而下降。随着欧盟WFD的实施,需要进一步减少营养负荷,这引起了贻贝渔民的关注,他们预计蓝贻贝的产量将进一步减少。
对这一复杂系统的管理采取了部门式的方法,对农业和减少富营养化有单独的治理和立法;渔业,包括贻贝捕捞;和自然保护。当农民等利益攸关方从具有高度环境影响的活动中受益,而他们本身并不直接依赖健康的海洋生态系统时,部门管理就不是最佳的。此外,在农业方面的规章制度和渔业方面的规章制度之间往往存在微妙的相互作用,这要求发展生物经济模型,将各部门之间的相互作用纳入其中,并对这些复杂系统提供更全面的表示和理解。
这项研究的目的是建立一个生物经济模型,对该系统进行综合描述,探讨在利姆峡湾建立生态和经济上可持续的贻贝渔业的管理方案。情景包括目前营养负荷的情况,以及根据欧盟世界营养日(http://ec.europa.eu/environment/water/water-framework/info/intro_en.htm).所检验的假设是:(1)实施世界水产养殖计划后,营养负荷的减少将导致贻贝产量的下降和贻贝渔业的利润减少;(2)贻贝渔业管理的改变将使贻贝渔业在实施世界水产养殖计划后仍能盈利。为了捕捉系统的复杂性,同时拥有一个可被涉众理解和接受的透明模型,并保持较短的计算时间,我们证明了在模型中结合机械过程描述和经验关系是一种有效的方法。
材料和方法
研究网站
利姆峡湾是一个浅的、富营养的峡湾系统,位于丹麦北部,有镶嵌的河口和海湾,向西连接北海,向东连接卡特加特(图1)。河口表面积约1526平方公里,集水区7528平方公里,农业占(62%)主要(Markager et al. 2006)。在过去的一百年里,总氮(N)和磷(P)的负荷增加了六倍。在20世纪80年代中期,最高的年装载量为13吨N/km²河口表层和0.91吨P/km²(Christiansen et al. 2006)。从那时起,养分负荷下降到8.2吨N/km²和0.30吨P/km²(kraouse - jensen等,2012年)。峡湾系统支持了大量底栖悬浮物的生物量(Maar等人,2010年),贻贝的收获主要基于对蓝贻贝自然种群的挖掘,贝壳类;只有少量的贡献来自于线上的贻贝养殖。自1995年以来,对利姆峡湾蓝贻贝渔业的管理一直是通过独家进入许可证,对船只的尺寸、发动机功率和配额以及关闭期限有明确的规定(Frost 2009年)。渔业的性质,单一的定居物种和单一的船队,加上独家进入许可证,满足了集体行动的要求(Ostrom 1991年)。对渔船的严格规格创造了一种同质结构,这虽然不是先决条件,但肯定有助于形成对渔业的集体管理行动。最初,这51个入境许可证是个人的,与船只捆绑在一起。然而,在2009年,许可证是可交易的,这导致船队减少到38艘,其中一些船有一个以上的许可证每艘船。每周配额、最小贻贝大小和适合钓鱼的贻贝肉的内容,以及食品安全法规管辖的渔业理事会(BEK没有。2000年7月3日第155号法律公告;2009年5月15日第372号https://www.retsinformation.dk/Forms/R0710.aspx?id=124647).但是,渔民们自愿集体减少了每艘船每周45吨的配额,这相当于水产厅规定的法定配额的50%左右。
Bioeconomic模型
生物经济模型是一个完全耦合的模型,由四个部分组成(图2)。它们是:(1)描述营养负荷与浮游植物初级产量之间关系的经验子模型;(2)基于过程的浮游植物和蓝贻贝生物量动态生态子模型;(3)基于代理的贻贝捕捞子模型,允许渔民根据法规、可用贻贝生物量、预期成本和收益来决定捕捞努力;(4)一个用于成本效益分析和模拟预期利润的经济子模型。生态模型组成部分基于Skive Fjord子盆地的监测数据;贻贝生产的经济数据涵盖了整个利姆峡湾(见图1),并从丹麦国民账户统计数据中获得。生态子模型由6个状态变量组成,一个为浮游植物生物量,5个描述蓝贻贝生物量组(图2)。浮游植物生物量以贻贝消耗的碳单位和叶绿素单位计算一个(排名一个)的浓度,以便与监测数据进行比较。贻贝生物量组为:M1,浅水贻贝(深度< 2 m);M2,长线生长的贻贝(水产养殖);在较深水区(> 2 m)有三个大小纲:M3,壳长< 20 mm的小个体(SL);M4,中等个人20-45毫米SL;M5,可收获贻贝,SL大于45 mm。
利用经验模型(Hinsby et al. 2012)建立了1月至6月和7月至12月两个独立时期N和P负荷与浮游植物初级产量之间的简化关系。这种半年一次的划分是基于以前的结果,结果表明磷在春季是主要的限制性营养物质,而氮在生长季节后期是限制性营养物质(Markager et al. 2006)。采用正向选择多元线性回归(MLR)方法(Sokal和Rohlf 2000)对1984-2003年丹麦监测项目(Markager等人,2008年)的数据建立了经验模型,包括初级产量和八个自变量中的一个选择:N负荷、P负荷、淡水径流、海水温度、全球表面辐射、NAO指数、盐度和风速³,其中N和P负荷作为强制变量加入MLR模型。NAO(北大西洋振荡)指数描述了冰岛和亚速尔群岛之间的压力差,它决定了主要的风型,从而决定了大规模的气候变化。
在生物经济模型中,浮游植物生物量(PB)被形容为
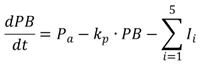 |
(1) |
浮游植物的摄食我这组贻贝依赖于温度和浮游植物浓度,被描述为
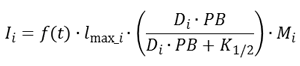 |
(2) |
研究区贻贝生物量我这取决于招聘人数、增长率和死亡率。生长速度是由食物的摄取量、排泄量和呼吸量决定的。死亡率包括自然捕食、缺氧和捕获引起的死亡率,因此取决于贻贝的深度和大小。在深水中的贻贝经常受到缺氧的影响,而在浅水中的贻贝很少经历缺氧的时期。海岸蟹(Carcinus maenas, L)及海星(阿斯忒瑞亚鲁本斯)是利姆峡湾主要的贻贝捕食者,因此浅水和深水贻贝的捕食压力都包括在内。在利姆峡湾,浅水是不允许进行疏浚的,只有> 45毫米长的贻贝被允许在较深的水域捕捞。因此,在模型中,只有最大尺寸类的贻贝(可捕捞贻贝)受到疏浚死亡率的影响。植物的生物量我贻贝群为:
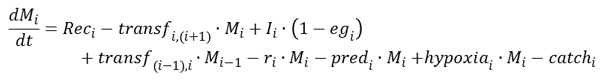 |
(3) |
如果一个地区是开放渔业,而贻贝的收获是有利可图的,预期的努力(E)被确定为用于捕获配额的努力或最大努力的最小值。
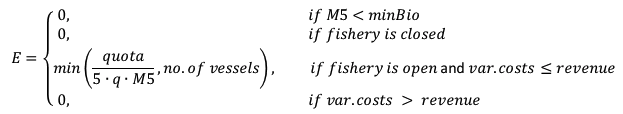 |
(4) |
除与实际捕捞作业有关的成本外,总捕捞成本亦包括固定成本(cf)与购买和维护渔船的费用有关。总成本为:
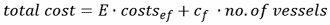 |
(5) |
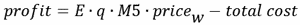 |
(6) |
生物经济模型中使用的所有生态经济参数值如表1所示。贻贝的招募、摄食、排泄和呼吸速率的生态参数值是根据文献值确定的(Vanharen和Kooijman 1993, Blackford 1997),其余参数值(捕食率和死亡率)按照下面的后验模拟进行校准。经济分模型中使用的参数是根据丹麦国民核算渔业生产统计数据统计估计的。在2000-2006年期间,挑选了11艘符合Limfjord渔场尺寸标准的船只。这些船只并不是每年都出现;样本中总共有66个观察结果。数据用于计算固定成本(cf)、可变成本(var.成本)和着陆价格(p).上岸后卖给贻贝产业,为世界市场生产产品。因此假定着陆价格反映了世界市场的情况。因此,由于Limfjord的产量只占全球供应量的一小部分,因此人们认为它不会影响价格。利用仿真软件ExtendSim v.7.1.5对方程进行数值求解。积分是用欧拉与每日时间步执行,这被证明是足够的数值精度所需的。
追算模拟
该模型从1984年到2003年校准了19年。在此期间,实施了几个国家水行动计划,导致包括利姆峡湾在内的所有丹麦河口的养分负荷显著减少(Krause-Jensen et al. 2012)。用于后向模拟的强迫数据(营养负荷、温度、风速、氧浓度等)从国家海洋数据数据库(MADS)程序(http://www.dmu.dk/en/-water/marinemonitoring/mads/)和丹麦气象研究所。通过对浮游植物和贻贝生物量模型模拟数据和观测数据之间的整体匹配,对浮游植物和贻贝死亡率参数值进行了仔细和系统的调整。散射图和R²值(Fitzpatrick 2009)用于最小化系统偏差和优化模型模拟和观测数据之间的拟合优度。反演模拟的结果在整个时期平均,Chl一个将浅层(深度≤2 m,第1组)和较深层(深度为> 2 m,第2 ~ 4组之和)水体的浓度、贻贝生物量与同期监测数据的年平均值进行比较。每周的背影一个从Skive Fjord的监测站获得了过去30年的数据,用于模型验证。Skive峡湾的贻贝生物量是根据潜水员对沿样带贻贝覆盖情况的观察,结合1993年至2003年期间25个监测站在深度区间0-2、2-4和4-6米采集的岩心样本(0.25 m²)估计的。
敏感性测试
通过对关键生态经济参数进行敏感性试验,确定模型对参数值选择的敏感性。在进行试验时,将选定参数的值分别改变1%、5%、20%和50%,保持所有其他参数不变并强制(营养加载),与后向模拟一样。场景
粮食日的实施预计将导致营养负荷的显著减少。2011年,通过了立法措施,预计氮的排放量将减少15%,并建议在2015年之后再减少17%。据估计,要达到《水框架指令》(Water Framework Directive)所要求的利姆峡湾河口良好的生态状态,1984年至2003年的平均载水量必须减少约47% (Markager et al. 2006)。这种显著的减少可能会影响整个生态系统,包括贻贝生产和贻贝疏浚业。为了探索未来养分负荷减少的生态和经济后果,采用与后置模拟相同的强迫值执行了WFD场景,但营养负荷相对于后置模拟中使用的负荷(WFD目标值)为47%。此外,通过改变每周配额、渔船总数和每艘渔船的捕鱼许可证数量,实现了贻贝渔业最优管理的场景,强迫和其余参数与后向模拟相同。结果
初级产量与养分负荷之间的经验关系
多元线性回归(MLR)模型描述春季(1-6月)浮游植物初级产量的结果如图3A和3B所示。解释变量包括磷负荷、盐度、风速³和地表辐射。MLR模型残差无系统性偏差,模型能描述73%的年际变化(图3B)。Skive Fjord的春季浮游植物产量从1984-2003年显著下降(图3A),与磷负荷的下降同步,磷负荷本身描述了50%的变化P一个为浮游植物产量。盐度是与北海水交换的代表,出现负系数,对应于河口磷浓度在与北海水交换高的年份被稀释。第三个变量是前一年7月至12月的平均风应力。这个变量对。有明显的负面影响P一个接下来的一年,可能是由于风应力和缺氧之间的负相关,这增加了沉积物中磷的泄漏。因此,秋天更多的风降低了第二年春天浮游植物可利用的磷浓度。选定的第四个也是最后一个变量是前一年7月至12月的平均地表辐射。高辐射降低了P一个第二年,可能由于地表辐射与植物秋季吸收的养分之间存在正相关关系,可用于来年春季初级生产的磷含量下降。在春季,磷负荷的系数高于其他解释变量,为0.82,盐度为0.096,风为-0.52,辐射为-0.15,证实了春季磷负荷的显著影响。这些系数可以直接比较,因为所有变量都被缩放为0的平均值和1的标准差。秋季(7-12个月)的浮游植物产量随时间变化较为稳定(图3C;1991年被认为是一个异常值),与春季初级产量相比。秋季初级生产的最终MLR模型有三个变量;N载荷、NAO指数、风速³(图3C, 3D)。N负荷的正向效应与预期一致,但显著低于春季P负荷的系数,也低于其他两个解释变量(N系数= 0.26,风速³-0.38,NAO指数-0.69)。模型中NAO指数和风应力的负面影响与春季盐度(水交换和稀释的代理)的负面影响以及风对缺氧的负面影响相对应。虽然秋季浮游植物产量的MLR模型与实测数据之间具有较好的相关性(R²= 0.42),但该模型往往低估初级产量的小值,高估大值(图3D)。
拥有强大的模式的影响P载荷在春天和N的弱效应加载在秋天也被观察到其他丹麦河口和其他变量比初级生产(2010年Markager et al . 2008年),可能反映了淡水载荷直接控制养分浓度出现在春天,而养分浓度季节更晚些时候由内部流程控制的河口,如缺氧的发生。
追算模拟1984 - 2003
以高Chl为特征的Skive峡湾浮游植物年动态一个模型充分再现了春季和夏季的浓度以及秋季较低的浓度(图4A)。年际变化,Chl偏高一个该模型也很好地描述了模拟期(1994-1988年)初期的浓度和其余时期较低的浓度。在20世纪90年代中期,四高氯化物一个记录了模型没有捕捉到的峰值。在浅水(深度≤2 m)和较深(深度为> 2 m)水域,观测到的贻贝生物量和模拟的年平均值具有良好的一致性(图4B, 4C)。这表明该生物经济模型对生态系统有很好的表征。然而,很难测试模型的因为Skive Fjord的贻贝生物量数据只提供了后6年的后6年。对无约束参数(如浮游植物的死亡率和贻贝的自然捕食率)的敏感性分析表明,模型对这些参数的较大变化(即≥20%)非常敏感,而且响应往往在食物链中向上传播,对浮游植物生物量的响应最低,而对利润的响应最高(图5)。此外,敏感性测试显示出很强的非线性。例如,深海贻贝捕食压力的降低导致贻贝生物量的增加,而登陆量和利润几乎不受影响(图5)。这表明,海星等的捕食在很大程度上控制着贻贝生物量,而在后发情况下的登陆量和利润则受到配额的控制。相比之下,当捕食压力增加50%时,贻贝的生物量和登陆量将分别减少75%和100%,表明渔民与其他贻贝捕食者之间存在竞争。对经济参数(如固定和可变成本和销售价格)的敏感性分析表明,在测试范围内,着陆不受经济参数变化的影响,但利润对参数值的变化呈线性反应。结果的场景
根据模型结果,将营养负荷降低到实施WFD的预期目标水平,即1989年至2003年平均负荷的47%,对浅水和深水贻贝生物量、登陆量和利润都有显著影响(图6A-D)。配额的减少表明,45吨/周/船的配额将优化登陆和利润在目前的营养负荷水平(图6D),并将确保较高的贻贝生物量在更深的水域(图6B)。达到WFD目标水平后,最佳经济配额估计为每艘船每周约30吨。然而,即使有一个最佳的捕捞配额,在实施WFD后,贻贝渔业也不会有利可图(图6D),因为贻贝产量和生物量减少,特别是在更深的水域(图6A, 6B)。以45吨/周/船的固定配额探索贻贝渔船数量的影响的情景(图6E-H)显示,在目前的营养负荷水平下,经济上的最佳渔船数量将是25艘。在过去的几年里,Limfjord的船只数量从51艘减少到38艘,但根据模型结果,应该进一步减少船只数量以优化利润。船只数量的减少将降低开采水平,导致在水深为> 2米的水域贻贝生物量增加三倍(图6F),而对食物(浮游植物)的竞争增加将使浅水贻贝生物量减少约50%(图6E)。如果贻贝的数量减少到能够达到世界粮食日指标的水平,那么贻贝的食物数量就会受到限制,生产力就会因此降低。贻贝产量的减少,意味着在实施野生渔护计划后,最理想的渔护船数目为约20艘(图7)。如果渔护船数目超过20艘,蓝贻贝的数量就会被过度捕捞,减少至最低水平(图6F)。在营养减少到WFD目标水平后,保持51个单个许可证被证明是不盈利的,即使在30吨/周/船的最佳配额(图7)。然而,如果目前的许可证数量分配到37艘船,配额至少为20吨/周/许可证,贻贝渔业将是盈利的。如果共享51个许可证的船只更少,贻贝渔业将在更低的配额下盈利。一个约30吨/周/许可证的配额优化利润,无论船舶数量。
讨论
海岸带的管理是一项挑战,需要基于科学的知识,因为生态过程及其与社会经济系统相互作用的复杂性(2011年诺贝尔奖)。由于缺乏适当规模的数据、缺乏跨部门和科学学科的交流以及缺乏对系统复杂性的理解等障碍,生物经济学模型和其他包含多个问题的跨部门模型非常少。此外,如果每个学科中的所有子学科都用最先进的模型表示,那么多学科模型将变得非常复杂。因此,与每个学科的现有标准相比,使用多学科模型需要灵活性和模型子部分的一些简化。这里提出的研究展示了第一个生物经济模型,该模型解决了与实现世界渔业发展目标有关的多个政策问题,以保持良好的生态状态,同时在丹麦海洋沿海系统维持有利可图的贻贝渔业。该模型是为Skive Fjord开发的,但也适用于其他有数据的地区。一个创新的方面是使用经验模型,通过在养分负荷和初级产量之间建立基于经验的联系来简化生物地球化学循环的描述(图3)。与流体动力学和生物地球化学循环的完全机制描述相比,使用经验捷径使模型透明,并确保较短的计算时间(CPU ~ 20秒/20年模拟)。这对于旨在促进利益相关者参与、利益相关者和科学家之间的讨论以及将数据与经济联系起来的模型至关重要。虽然经验子模型不像机械描述那样通用,但该模型包含了控制初级产量的特征特征,如盐度和风(即水动力学)、营养负荷和季节性。因此,当有必要的长期监测数据时,无论是单独还是与动态模型结合,经验联系都是有用的,如本研究中所做的那样。模型验证是测试模型可信度和增加模型场景置信度的重要工具(Fitzpatrick 2009)。不幸的是,由于数据可用性的限制以及人力和技术资源的限制,在集成模型中,模型结果与经验数据的数值比较验证常常被忽略(van Delden et al. 2007)。后验模型运行表明,生物经济模型能够重现20年的观察到的Chl的时间序列一个数据,包括Chl的下降趋势一个模拟期间观察到的浓度(图4A)。20世纪80年代中期,高(> 40µg L-1)的背影一个浓度定期发生。尽管该模型没有捕捉到这些峰值的确切时间,但其规模很好地反映了出来。减少的背影一个尤其与磷含量的显著降低相吻合。养分负荷与Chl之间存在显著关系一个的变化,因为内部负荷和浮游动物或贻贝的捕食等因素的变化,以及Chl一个降低营养负荷的浓度通常很低(Timmermann等,2010年)。四个背影一个该模型没有捕捉到上世纪90年代中期记录的峰值。这些峰值出现在8月或9月,极有可能是由于缺氧沉积物中的营养物质释放造成的(Carstensen et al. 2007)。这表明了经验模型的局限性,它可以再现年际变化的总体模式,但不能捕捉特定事件。一个动态模型,包括底栖营养循环,可能会更好地捕捉具体事件,但它也会使模型复杂、缓慢和不透明,并会阻碍利益相关者的交互使用。在后验期观测到的强迫(养分负荷)变化较大,模型结果与经验数据的一致性普遍较好(图4),说明模型不仅能够再现数据,而且对强迫的变化也能做出正确的响应,这对情景可靠性至关重要。此外,较长的模拟周期表明,累积误差和错误趋势没有出现。
观测到的贻贝生物量与模拟的贻贝生物量之间存在合理的一致性(图4B, 4C)。该模型似乎捕捉到了正确的生物量水平,但观测到的生物量的年变化似乎比模型预测的要高。此外,该模型较好地反映了浅水和深水贻贝生物量的相对分布。
在丹麦,世界生态日的实施是由环境治理推动的,其目的是重建良好的生态状况,而资源利用的管理主要是为了符合环境目标。这种碎片化的管理可能无法在不断变化的压力和基线下实现自然保护和资源利用之间的可持续平衡。在利姆峡湾,贻贝渔业是通过定期的环境影响评估进行管理的,该评估考虑贻贝生物量的承载能力,而不考虑经济后果。作为对这种管理的回应,贻贝渔民之间达成了一项公共协议,将当局设定的配额从85吨/船/周减少到45吨,他们认为这样在经济上更可行(Dinesen et al. 2011)。生物经济模型的结果证实了可持续贻贝渔业的利益攸关方共识,该模型显示,在这种捕捞强度下,利润是最佳的。利益攸关方的共识是基于渔民的集体经验,这些经验是通过探索性捕捞获得贻贝数量、大小和肉含量的信息。虽然船队的规模已从51艘减少到38艘,但与目前的模式相比,渔民未能通过集体协议达到25艘的最佳船队规模。有三个原因可以解释这一点:(1)协议和行动不是建立在现有模式的基础上,它们是在缺乏知情的基础上发展起来的;(2)船舶减少是通过贸易,需要一段时间;(3)牡蛎许可证也可以交易,一些船只同时拥有牡蛎和贻贝许可证。
随着条件的变化,如营养负荷的减少和贻贝生物量的减少,渔民需要应对资源的进一步减少。特别是在这种变化期间,渔民和环境将受益于能够预测资源可用性的主动管理,从而支持可持续利用,同时优化利润并努力满足自然保护的要求。
在这项研究中,生物经济模型被证明能够预测系统变化的生态和社会经济后果,包括环境压力。只要生态系统的状态在模型数据支持的范围内,预测就是可靠的。由于预期的营养负荷下降,贻贝渔业管理结构必须从船只配额制度转变为允许船只拥有多艘许可证的许可证制度(图7)。维持51艘船只的单一配额在经济上是不可行的,因为预计贻贝存量和渔获量会下降。利润优化建议减少到20艘船,这将优化贻贝渔民的利润。然而,如果贻贝生物量减少到最低水平,则需要进一步减少到10艘,以支持可持续的资源利用(图6E, 6F)。根据现行的许可证制度,这意味着允许拥有两个以上许可证的船舶减少,每个许可证为30吨/周。在实践中,这意味着更少的渔民参与这个行业,而挑战将是如何在更少的参与者中分配许可证。然而,模型结果允许管理人员在这方面积极主动,这可以促进一个成功的结果。
营养负荷和底栖生物产量之间的相互作用是复杂的,其结果取决于食物供应和缺氧之间的平衡(Timmermann et al. 2012)。尽管我们提出的模型模拟表明,利姆峡湾的贻贝产业明显受益于富营养化,但众所周知,富营养化和缺氧会损害其他渔业产业(Huang和Smith 2011),并带来严重的经济后果(Huang et al. 2012),而且,由于营养物质减少,利姆峡湾的其他生态系统服务很可能会增加。一种可能的情况是,在20世纪80年代和90年代初,由于营养负荷的负面影响,鲽鱼和比目鱼等固定的鳍鱼数量急剧减少(Christiansen et al. 2006),将返回河口,并支持当地的渔业。鳝鱼草的重新定居是未来营养负荷减少的另一个预期结果;这将对鳗草草甸的生态系统服务产生显著的积极影响,包括营养固定、沉积物稳定性和碳储存(Duffy 2006, fourqureen et al. 2012)。减少营养负荷的这些和其他潜在的积极影响,例如对休闲渔业和旅游业的影响,没有包括在本模型中,因此该模型仅限于预测贻贝捕鱼业的社会经济后果,并探索在不断变化的环境压力下获得经济、可持续贻贝渔业的潜在管理策略。SAF的优势在于生态和社会经济组成部分的耦合,包括与系统的生态和社会经济过程相关的利益相关方的知识和经验以及经验数据。
SAF方法是通用的,已经在欧洲各地的沿海地区进行了测试(Hopkins et al. 2012)。例如,Vermaat等人(2012)开发了一个空间动态模型,允许通过考虑当前和未来的社会环境来改善农业决策。Melaku Canu等人(2011)和Schernewski等人(2012)都演示了如何使用SAF来解决可持续双壳贝类养殖和减少非法捕捞。有了SAF,威尼斯泻湖的大规模蛤蜊养殖可以更加可持续,可以为当地社区实现社会和经济效益,同时减少负面的环境影响和社会冲突(Melaku Canu et al. 2011)。同样,通过利益攸关方的参与,可以改善Thermaikos湾的小规模贻贝生产;他们对综合管理工具的理解得到了提高,并提供了调查场景(Konstantinou等,2012)。多学科科学、利益攸关方参与和使用模型情景的结合使SAF方法非常适合解决需要多学科和整体分析和管理战略的政策问题。例如,在切萨皮克湾(Chesapeake Bay),非法捕捞牡蛎对生态系统产生了负面影响,使恢复牡蛎数量及其恢复力的努力成为徒劳(Bashore et al. 2012)。SAF方法可用于让所有利益攸关方参与进来,并展示当地个人如何从基于探索场景的综合管理工具中受益,其方式与Melaku Canu等人(2011)和Konstantinou等人(2012)的方法类似。SAF方法(Hopkins et al. 2011, 2012)为真实世界提供了可识别的、透明的描述,并允许通过生物经济模型模拟更好地理解系统复杂性。 It engages stakeholders from the outset, thereby ensuring ownership and facilitating self-policing and higher acceptance of management regulations.
结论
利用生物经济模型进行的模型模拟显示,在实施WFD后,二次生产大幅减少,这将导致Limfjord河口贻贝渔业的崩溃。然而,在未来生产力较低的环境中,通过将渔船数量减少到大约10艘,并允许每艘渔船获得多个许可证,贻贝渔业可以在生态和经济上可持续发展。利用经验模型简化复杂的生物地球化学循环,使模型透明,计算时间短,这对于旨在促进利益相关者参与的模型至关重要。
致谢
本研究部分由欧盟委员会的欧盟第6框架计划(合同编号036992)资助;由丹麦食品工业机构(DFFE)资助的林姆峡湾国家制度转变项目;由国家项目丹麦河口的营养负荷和环境质量,也由DFFE和MarBioShell项目,由丹麦科学、技术和创新机构资助。我们感谢奥胡斯大学Morten Hjort在模型开发期间提供的技术援助。此外,我们还要感谢高级顾问研究员埃里克·霍夫曼、Per Dolmer和Per S. Kristensen;导航系统Aqua Charlottenlund;和SPICOSA Limfjord利益相关者小组,以了解Limfjord贻贝生产的信息和讨论。我们非常感谢那不勒斯大学的汤姆·霍普金斯教授对手稿提出的宝贵意见。
文献引用
巴肖尔,c.j., H. A.莱恩,K. T.佩恩特,M.内勒,J. R.哈丁,D. C.拉夫。2012。海警因非法捕捞东区生蚝而被传讯及司法裁决的分析(Crassostrea virginica,从1959年到2010年,在美国切萨皮克湾的马里兰州部分发现了Gmelin 1791)。贝类研究杂志31:591 - 598。http://dx.doi.org/10.2983/035.031.0301布莱克福德,j.c. 1997。北海生态系统模型底栖生物动力学分析。海洋研究杂志38:213 - 230。http://dx.doi.org/10.1016/s1385 - 1101 (97) 00044 - 0
Carstensen, J. P. Henriksen和A. S. Heiskanen. 2007。浅水河口夏季藻华:定义、机制和与富营养化的联系。湖沼学和海洋学52:370 - 384。http://dx.doi.org/10.4319/lo.2007.52.1.0370
克里斯汀森,T. J.克里斯汀森,S. S.马克格,J. K.彼得森和L. T.莫里特森。2006。Limfjorden i 100 år。Klima, hydrografi, næringsstoftilførsel, bund动物群og fisk i Limfjorden, 1897年至2003年。578年的报告。丹麦Miljø在20ø凝胶。和DMU的关系很融洽。国家环境研究所,罗斯基勒,丹麦。(在线)网址:http://www2.dmu.dk/1_viden/2_Publikationer/3_fagrapporter/rapporter/FR578.pdf
迪内森,g.e., K. Timmermann, E. Roth, S. Markager, L. Ravn-Jonsen, M. Hjorth, M. Holmer, J. G. Støttrup。2011.丹麦Limfjord的贻贝生产和水框架指令目标:用于系统管理的综合评估。生态和社会16(4): 26。http://dx.doi.org/10.5751/ES-04259-160426
多尔默,P. R. P.弗兰森,2002。丹麦贻贝渔业的评价:生态系统管理方法的建议。Helgoland海洋研究56:13-20。http://dx.doi.org/10.1007/s10152-001-0095-6
达菲,2006。生物多样性和海草生态系统的功能。《海洋生态发展系列311:233 - 250。http://dx.doi.org/10.3354/meps311233
Engelen, G. I. Uljee和K. Van de Ven, 2003。WadBOS:整合知识,支持瓦登海的政策制定。513 - 537页在S.格特曼和J.史迪威,编辑。在实践中规划支持系统。施普林格,柏林,海德堡,德国。http://dx.doi.org/10.1007/978-3-540-24795-1_28
菲茨帕特里克,2009年。水质管理人员对河口和海岸富营养化模型的评估技能。海洋系统学报76:195 - 211。http://dx.doi.org/10.1016/j.jmarsys.2008.05.018
Fourqurean, J. W., C. M. Duarte, H. Kennedy, N. Marba, M. Holmer, M. A. Mateo, E. T. Apostolaki, G. A. Kendrick, D. kraus - jensen, K. McGlathery, O. Serrano. 2012。海草生态系统是全球重要的碳储备。自然地球科学5:505 - 509。http://dx.doi.org/10.1038/ngeo1477。
弗罗斯特,H. S.安德森,M.尼尔森。2009,Økonomisk tilpasning直到miljømål i blåmuslingefiskeriet。国家økonomisk tidsskrift147(2): 158 - 174。
加西亚,S. M.和K. L.科克伦,2005。渔业的生态系统方法:执行准则的审查。国际内燃机学会海洋科学杂志62:311 - 318。http://dx.doi.org/10.1016/j.icesjms.2004.12.003
格特曼和J.史迪威,编辑。2009.规划支持系统最佳实践和新方法.施普林格。http://dx.doi.org/10.1007/978-1-4020-8952-7
辛比,K., S. Markager, B. Kronvang, J. Windolf, T. O. Sonnenborg, L. Thorling. 2012。生态状况较差的温带河口集水区营养物的阈值及管理方案。水文与地球系统科学“,16:2663 - 2683。http://dx.doi.org/10.5194/hess-16-2663-2012
霍普金斯,T. S., D.贝利,R.埃尔姆格伦,G.格莱格,A.桑德伯格,J. G. Støttrup。2012.向可持续发展过渡的系统方法框架:基于沿海试验的潜在价值。生态和社会17(3): 39。http://dx.doi.org/10.5751/ES-05266-170339
霍普金斯,T. S. D.拜利,J. G. Støttrup。2011.沿海地区的系统方法框架。生态和社会16(4): 25。http://dx.doi.org/10.5751/ES-04553-160425
黄l ., L. A. B.尼科尔斯,J. K.克雷格,M. D.史密斯。2012。衡量因缺氧造成的福利损失:以北卡罗莱纳棕虾为例。海洋资源经济学27:3-23。http://dx.doi.org/10.5950/0738-1360-27.1.3
黄,L.和M. D.史密斯,2011。存在生态压力的年度渔业管理:虾和缺氧的情况。生态经济学70:688 - 697。http://dx.doi.org/10.1016/j.ecolecon.2010.11.003
伊梅森,J. C. J. M.范登伯格,2004。贝类渔业的生物经济学分析:元种群模型中招募和生境的影响。环境与资源经济学27:65 - 86。http://dx.doi.org/10.1023/B:EARE.0000016797.30911.3f
Konstantinou, Z. I., Y. N. Krestenitis, D. Latinopoulos, K. Pagou, S. Galinou-Mitsoudi,和Y. Savvidis. 2012。希腊Thermaikos海湾Chalastra的贻贝养殖活动:解开管理难题的努力。生态和社会17(1): 1。http://dx.doi.org/10.5751/ES-04455-170101
Krause-Jensen, D. S. Markager和T. Dalsgaard. 2012。底栖和远洋生物在不同营养条件下的初级生产。河口和海岸35:527 - 545。http://dx.doi.org/10.1007/s12237-011-9443-1
马尔,M., K. Timmermann, J. K. Petersen, K. E. Gustafsson, L. M. Storm, 2010。利姆峡湾(Limfjorden)浅水口蓝贻贝营养负荷和水柱稳定性调节的模型研究。海洋研究杂志64:322 - 333。http://dx.doi.org/10.1016/j.seares.2010.04.007
Markager, S. M.巴松皮埃尔和D. L. J.彼得森,2008。分析1986年至2006年的miljøtilstanden和Mariager峡湾。miljøtilstanden经验主义建模。685年的报告。丹麦奥胡斯大学国家环境研究所。(在线)网址:http://www2.dmu.dk/Pub/FR685.pdf
马克格斯,卡斯滕森,克劳斯-詹森,温道夫,蒂默曼,2010。Effekter af øgede kvælstoftilførsler på miljø i danske峡湾。报告787,奥尔胡斯大学国家环境研究所,奥尔胡斯,丹麦。(在线)网址:http://www2.dmu.dk/Pub/FR787.pdf
斯道姆(c.a. Stedmon)。2006。Limfjordens miljøtilstand 1985 - 2003。Sammenhæng mellem næringsstoftilførsler, klima og hydrografi相信经验模型。报告577,国家环境研究所,罗斯基勒,丹麦。
Melaku Canu, D, P. Campostrini, S. Dalla Riva, R. Pastres, L. Pizzo, L. Rossetto和C. Solidoro. 2011。解决威尼斯泻湖蛤蜊养殖的可持续性问题。生态和社会16(3): 26。http://dx.doi.org/10.5751/ES-04263-160326
诺布雷,2011年。用科学的方法解决沿海管理的挑战。《海洋生态发展系列434:279 - 289。http://dx.doi.org/10.3354/meps09250
奥斯特罗姆e . 1991。治理公地:集体行动制度的演变.剑桥大学出版社,英国剑桥。http://dx.doi.org/10.1017/CBO9780511807763
Schernewski, G. N. Stybel和T. Neumann. 2012。Szczecin (Oder)泻湖的斑马贻贝养殖:水质目标和成本效益。生态和社会17(2): 4。http://dx.doi.org/10.5751/ES-04644-170204
索卡尔,R. R.和F. J. Rohlf. 2000。生物计量学:生物学研究中统计学的原理和实践.第三版。W. H.弗里曼,纽约,美国纽约。
泰特,P, A.桑德伯格,A.梅特,编辑。2011.持续的海岸带系统.达尼丁文献出版社,德文郡,英国。
Timmermann, K. S. Markager, K. E. Gustafsson, 2010。小溪还是大海?用三维水动力-生态模型追踪浅水河口营养盐的来源和影响。海洋系统学报82:111 - 121。http://dx.doi.org/10.1016/j.jmarsys.2010.04.008
Timmermann, K. J. Norkko, U. Janas, A. Norkko, B. G. Gustafsson, E. Bonsdorff. 2012。模拟大型动物群生物量与缺氧和营养负荷的关系。海洋系统学报105:60 - 69。http://dx.doi.org/10.1016/j.jmarsys.2012.06.001
Tomczak, M., G. E. Dinesen, E. Hoffmann, M. Maar, J. G. Støttrup。2013.林姆峡湾(丹麦)生态系统变化的综合趋势评估:最近政权转移的证据?河口海岸和陆架科学117:178 - 187。http://dx.doi.org/10.1016/j.ecss.2012.11.009
Tserpes, G., E. Tzanatos, P. Peristeraki, V. Placenti和L. Kell. 2009。地中海剑鱼不同管理措施的生物经济评价。渔业研究96:160 - 166。http://dx.doi.org/10.1016/j.fishres.2008.10.008
范德尔登,H., P. Luja和G. Engelen. 2007。流域管理中社会经济和物理过程多尺度动态空间模型的集成。环境建模与软件22:223 - 238。http://dx.doi.org/10.1016/j.envsoft.2005.07.019
范哈伦,R. J. F.和S. A. L. M. Kooijman. 1993。动态能量预算模型的应用贝壳类(左)。荷兰海洋研究杂志31:119 - 133。http://dx.doi.org/10.1016/0077 - 7579 (93) 90002 -
Vermaat, J. E. S. Broekx, B. Van Eck, G. Engelen, F. Hellmann, J. L. De Kok, H. Van der Kwast, J. Maes, W. Salomons和W. Van Deursen. 2012。Scheldt河集水区、河口和邻近沿海水域氮源分配。生态和社会17(2): 30。http://dx.doi.org/10.5751/ES-04889-170230
