

去pdf本文的版本
De la Monta / a, E., R. del Pilar Moreno-S / nchez, J. H. Maldonado和D. M. Griffith. 2015。预测厄瓜多尔亚马逊土著社区的猎人行为:来自家庭生产模型的见解。生态和社会20(4): 30。
http://dx.doi.org/10.5751/ES-08032-200430
预测厄瓜多尔亚马逊土著社区的猎人行为:来自家庭生产模型的见解
摘要
许多生活在亚马逊的土著社区依靠狩猎和捕鱼来满足他们对蛋白质的大部分需求。尽管这些做法很重要,但该地区很少有研究从家庭层面分析打猎和捕鱼的社会经济驱动因素。我们提出了一个家庭生产模型,以评估位于厄瓜多尔亚马逊地区的小型土著社区的关键经济参数对狩猎和渔业的影响,该社区的蛋白质主要来源来自狩猎和渔业。该模型使用来自两个反映不同市场整合和森林保护水平的社区的经验数据进行了验证。需求和供给函数是根据19个月来收集的家庭数据生成的。推导出弹性,以确定从事狩猎决策对外生参数的敏感性,如非农工资、狩猎成本、丛林肉价格、对非法销售丛林肉的惩罚和猎物物种的生物学特征。在对模型进行校正后,我们模拟了关键经济参数的变化。最直接影响两个社区狩猎活动的参数是非农工资。模拟10%的工资增长会导致收获生物量减少16-20%,而50%的工资增长会使收获生物量减少> 50%。模型模拟显示,野生动物肉的价格和对非法贸易的惩罚也对猎人的行为有重要影响,就野生动物肉的销售数量而言,但对总收获量而言没有影响。 As a tool for understanding hunters’ economic decision-making, the model provides a basis for developing strategies that promote sustainable hunting and wildlife conservation while protecting indigenous livelihoods.介绍
野味是蛋白质的主要来源,也是发展中国家数百万居民的重要收入来源。每年,野生动物肉的收获、销售和消费价值数十亿美元(Milner-Gulland et al. 2003),有人认为,如果可持续管理并有效地纳入整体经济,这些活动可以为发展做出积极贡献(Brown 2003)。然而,尽管在大多数热带地区,狩猎对家庭经济的重要性(Bodmer et al. 2004, de Merode et al. 2004),不可持续的丛林肉开发构成了对当地生物多样性的主要威胁之一(Vié et al. 2009)。
目前亚马逊地区大部分地区的经济和社会状况,加上那些提供和消费丛林肉的人的文化特征,意味着对于大多数物种来说,狩猎是不可持续的(Milner-Gulland et al. 2003)。狩猎不仅威胁到物种的灭绝,还可能对生态系统及其提供的商品和服务产生严重影响(Fa et al. 2002)。这反过来又威胁到直接依靠自然生态系统运作维持生计的土著人民的粮食安全和文化完整。
在厄瓜多尔亚马逊地区,丛林肉的消费水平相对于其他亚马逊地区较高(Fa和Peres 2001, de la Montaña 2013),其结果是对生物多样性的负面影响,包括当地物种的灭绝,是经常发生的(Franzen 2006, Zapata-Ríos et al. 2009)。如果要避免这些影响,就必须通过改变习惯和实施确保可持续野生动物管理的战略来减少家庭消费和商业需求。
尽管狩猎对农村人口的重要性,以及过度开发对当地生计和生物多样性造成的危险,但影响家庭层面丛林肉消费的因素仍然知之甚少(Brashares et al. 2011)。大多数关于拉丁美洲丛林肉的研究都集中在了解狩猎实践和收获水平的可持续性上(Robinson和Redford 1991, Peres 2000, Sirén等2004,Levi等2009,Shepard等2012,Iwamura等2014)。少数以经济为重点的研究基于经验数据检验了丛林肉收获和家庭变量之间的关系(Wilkie和Godoy, 2001年,Demmer等人,2002年,Sirén等人,2006年,Godoy等人,2010年)。据我们所知,只有一项来自非洲的研究明确地将野生动物肉的开发作为家庭经济的组成部分(Damania et al. 2005)。然而,此前没有研究建立家庭层面的社会经济因素对拉丁美洲狩猎的影响模型。
我们开发了一个家庭生产模型,旨在确定小型土著社区狩猎和渔业活动最重要的社会经济驱动因素。我们用来自亚马逊地区两个代表不同社会经济现实的社区的数据验证了模型,并分析了关键经济参数对狩猎的影响,如非农工资、狩猎成本、丛林肉价格和对非法销售丛林肉的惩罚。模型结果为猎人的经济决策提供了洞见,并有助于确定促进可持续狩猎和野生动物保护的战略,同时保护土著居民的生计。
方法
研究领域
该研究是在位于厄瓜多尔东北部的Cuyabeno野生动物生产保护区内及其附近进行的(图1)。该地区形成了生物多样性热点的一部分(Myers et al. 2000, Brooks et al. 2006),是一个全球重要的生态区(Olson and Dinerstein 2002)。Cuyabeno保护区包含大约6000平方公里的热带雨林(Sierra 1999),有12000种植物和1320种脊椎动物,其中哺乳动物、鸟类和鱼类分别为165、493和475种(Rivadeneira 2007)。
居住在保护区内外的人口约有2000人,属于五个不同的民族(A 'i Cofán, Secoya, Siona, Kichwa, Shuar),以及居住在保护区外围的非土著殖民者(Instituto Nacional de Estadística y Censos 2010)。保护区的西半部被次生林、牧场、自给作物和油棕种植园包围。1964年在该地区开始的大规模石油开采作业最近已扩展到该保护区。
为了用反映该地区土著社区不同社会经济背景和狩猎动态的数据来验证模型,我们收集了来自A 'i Cofán和Secoya少数民族的两个土著社区的数据(表1)。这两个社区在规模和生存方式方面都是厄瓜多尔亚马逊地区的代表(de la Montaña 2013),但在市场准入和相对于保护区的位置方面存在差异(图1)。
Zábalo位于保护区内的大片成熟原始森林中,只有通过河流才能到达。这个社区远离城市中心,与市场经济的联系并不紧密。打猎和钓鱼是蛋白质的主要来源,而且经常进行。自给作物,如木薯、香蕉和其他水果,以及商业产品,如可可,都种植在面积小于1公顷的家庭土地上chacras。
Wajosará位于保护区外,被耕地、次生林和比Zábalo小得多的原始森林所包围。Wajosará距离大量人口中心不超过一小时的路程,可通过公路到达,靠近可以获得基本商品的小定居点。与Zábalo类似,维持生计的狩猎和捕鱼是日常活动,木薯和香蕉生长在chacras。2010年,一些Secoya家庭开始种植油棕。
的理论模型
为了了解关键经济参数对丛林肉提取的影响并模拟猎人行为,我们基于Damania等人(2005)提出的生物经济模型开发了一个家庭生产模型,并对其进行了调整,以反映所研究社区的具体特征。该模型包括该地区家庭进行的三种主要生产活动,即打猎、捕鱼和非农工作,并假定从这些活动中获得的收入用于购买粮食和专门供家庭使用的其他资源。
我们没有区分游戏物种,而是将所有物种都视为具有特定种群密度和捕获概率的单一代表性游戏物种。我们还假设,出售该物种是非法的,但可以提取以维持生计。该模型通过假设对狩猎活动进行监测,并且有一定概率发现出售狩猎所得肉类的家庭,从而纳入了这一限制。在这种情况下,野味被认为是被没收的。此外,该模型假设猎人不使用陷阱,而是积极地使用各种武器(火器、砍刀、狗等)追捕猎物,这是新热带土著社区的普遍情况(Fa和Peres 2001)。因此,我们只提出一个狩猎生产函数,它与猎人使用的武器无关。
家庭表现出柯布-道格拉斯型效用函数:
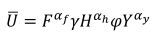 |
(1) |
在哪里F家庭消费的商品不是从打猎和捕鱼获得的产品(例如,其他蛋白质来源、食品或消费品);H是家庭捕获的动物的生物量;Y是家庭捕捞鱼类的生物量;αf+αh+αy= 1,表示商品(F)、野味(H)和鱼(Y)对家庭产生效用;γε[0,1]是家庭消费丛林肉的比例,而(1−γ)为售出的比例;而且φε[0,1]是家庭消费鱼类的比例,而(1−φ)为销售比例。
根据式1,给定家庭的效用取决于商品(F)、野味(H)和鱼(Y).渔猎劳动者家庭效用呈现正相关关系,随消费增加而递减。换句话说,效用随着消费的增加而增加,但以递减的速度增加。因此,
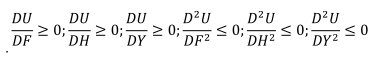 . . |
(2) |
家庭还面临一些约束,包括预算约束、狩猎生产约束、渔业生产约束和劳动力约束。预算约束由
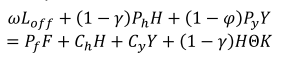 |
(3) |
在哪里ω是通过出售劳动力而获得的工资,l从是把劳动时间用在了非农工作上,Pr价格好吗r(r=f、h、y),Ch是狩猎投入的单位成本,Cy为捕鱼投入的单位成本,θ被发现贩卖野味的概率,和K是对出售野味的惩罚。在这种情况下,K相当于从狩猎中获得的没收产品的等值货币价值。对销售从捕鱼中获得的产品的情况不适用处罚。
因此,表达式ωL从表示出售劳动所得的家庭收入;(1−γ)PhH是卖野味获得的收入;(1−φ)PyY是卖鱼所得的收入;PfF代表家庭在其他消费品上的支出;ChH而且CyY分别表示打猎和捕鱼活动的投入费用;和(1−γ)K Hθ表示因出售野味而受处罚的预期货币价值。
狩猎生产函数根据Damania等人(2005)表示为
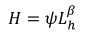 |
(4) |
在哪里ψ为猎物存量和捕获概率对收获生物量的影响;lh是把劳动的时间奉献给打猎;而且β是一个技术参数,表示致力于狩猎的劳动力的生产力,0 <β< 1。我们假设致力于狩猎的劳动力和丛林肉的收获之间存在非线性关系。
渔业生产函数定义为
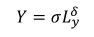 |
(5) |
在哪里σ是鱼类资源和捕获概率对收获生物量的影响;ly是把劳动的时间奉献给钓鱼;而且δ为技术参数,表示从事渔业劳动的生产率,0 <δ< 1。与狩猎一样,我们假设致力于捕鱼的劳动力与收获之间存在非线性关系。
劳动约束被定义为
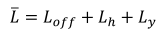 |
(6) |
在哪里L̑是用于打猎、钓鱼和其他创收活动的全部家庭劳动时间。根据这一限制,用于生产活动的时间分布在非农工作(l从)、专门从事狩猎的劳动(lh)和专门从事捕鱼的劳动力(ly).
猎人-渔民-工人家庭面临的经济问题是效用最大化,但受制于前面规定的限制:
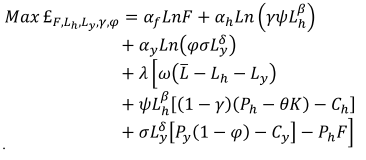 . . |
(7) |
这个优化问题的解决方案产生了狩猎(lh)和钓鱼(ly)及家庭对野味的需求(γH)、鱼(φY)及其他货物(F)作为参数的函数H, Y, L从,Ph,Py,Pf,Ch,Cy,ω,αf,αh,αy,β, δ, ψ, σ, θ而且K。为了进行比较静力学,我们估计了相关变量的弹性(H而且Y)至主要经济参数的变化。
实证模型
数据收集
在2013年2月至2014年8月的19个月期间,我们调查了35个同意参与研究的家庭,占两个社区家庭总数的90%(表1)。调查的9个月和10个月分别对应于旱季(12月- 2月和7月- 8月)和雨季(3月- 6月和10月- 11月)。我们设计并实施了四种类型的调查。首先,我们要求社区成员完成一项登记调查,要求了解每户家庭的基本信息。其次,我们与来自社区的10名现场助理合作,每周进行一次家庭调查,收集有关人口、人力资源、收入、投入成本、丛林肉和其他食品的价格、丛林肉的销售量以及家庭对丛林肉和其他动物蛋白质来源的消费量的信息。每名外地助理都接受了进行调查问题单的个别培训。第三,在同一时期内,现场助理每天都对家庭进行访问,以尽量减少由于受访者记忆错误而造成的数据丢失。要求家庭成员具体说明捕获动物的数量和种类、用于生产活动的时间、捕获物种的最终用途以及与其他家庭分享的数量。最后,我们要求两个社区的猎人在每次狩猎后完成一项调查。这项调查的目的是获得关于狩猎地点、捕猎努力和成本以及猎物的生物学方面的补充信息。为了方便非西班牙语使用者填写,我们提供了西班牙语和相关本土语言的问卷。 We also trained participants to weigh each animal caught using a set of scales.
模型的校正
为了用从每个群落收集的经验数据校准模型,我们计算了观察到的内生和外生变量的平均周得分(表2)。H)以准备好胴体后的猎物重量计算,而鱼类生物量(Y)与调查报告的渔获总重量相吻合,因为我们无法获得有关鱼的内脏重量的资料。未直接测量的参数(即,αf,αh,αy,ψ, σ, β, δ)使用理论模型导出的方程和最佳调整的数值逼近进行估计(表3)。我们使用这些参数估计来校准模型,建立观察到的内生变量与其基于估计的供给和需求函数的模型预测值之间的对应关系。
模拟
为了评估经济变量变化对狩猎的影响,我们使用估计的弹性和校准的模型模拟了不同的场景。我们通过逐个修改关键的经济参数(保持所有其他参数相同)来构建场景,并分析这些变化如何影响每个研究社区中有关狩猎的家庭行为。
结果
理论模型
供给和需求函数的估计
解决优化问题得到了渔猎劳动力供给作为参数的函数(见附录1)。A1.1和A1.2分别表明,可用于狩猎或捕鱼的家庭劳动力直接取决于:(1)野生动物肉或鱼类的价格,以及(2)增加野生动物或鱼类种群自然产量的生物因素。相比之下,专门用于狩猎或捕鱼的劳动力与与狩猎或捕鱼相关的成本、出售非农劳动力产生的工资以及(就狩猎劳动力而言)对非法销售野生动物肉的预期惩罚呈负相关。
该理论模型还使我们能够推导出对野味和鱼类的需求(见附录1)。A1.3和A1.4表明,对这些产品的需求直接取决于:(1)这些产品的消费为家庭提供的效用,以及(2)家庭收入,家庭收入是出售野生动物肉和鱼类以及打猎和捕鱼以外的活动所获得的预期净收入之和。相反,对野味和鱼类的需求则与这些商品的预期价格成反比。值得注意的是,出售野味的预期价格反映了家庭消费野味的隐含或影子价格。
弹性估计
为了评估供需对关键经济参数(工资、投入成本、产品价格、出售野味的惩罚成本等)变化的响应,我们通过估计兴趣变量的弹性来应用比较静力学:H)和钓鱼(Y;附录2,表A2.1)。例如,正如理论预测的那样,丛林肉和鱼的供应弹性与价格呈正相关。丛林肉或鱼类价格的上涨分别提高了家庭从事的狩猎或捕鱼的水平。
经验模型
描述性统计
狩猎为两个社区的家庭提供了最重要的蛋白质来源,也就是说,比家庭或商业来源更多。毫不奇怪,考虑到更多的家庭数量,Zábalo的居民在研究过程中比Wajosará的居民猎杀更多的动物:2128只动物代表14164公斤,而416只动物代表3138公斤。Zábalo的家庭平均体重为38.1公斤,也较高−1月−1相比之下,家庭体重20.2公斤−1月−1在Wajosara。渔业在两个社区的饮食中也扮演了重要的角色。与丛林肉类似,Zábalo的鱼类总消费量高于Wajosará:分别为5004公斤和1963公斤。但是,每户13.2公斤和12.9公斤的消费量大致相同−1月−1分别在Zábalo和Wajosará。
丛林肉和鱼类的消费模式在两个群落中都表现出季节性,这可能是物种自然循环的结果(图2、3)。在Zábalo中,有证据表明鱼类和丛林肉的消费之间存在互补性,即一种资源的低可用性时期通过对另一种资源的更高开发得到补偿,导致总体消费比率相当恒定(图2)。
收入来源的比较显示,Zábalo的生产活动比Wajosará的生产活动更以生存为基础,这从Zábalo的生产活动和采掘活动的总收入较低可以看出(图4和图5)在Zábalo,平均每户家庭每周26.8美元的劳动几乎是家庭收入的唯一来源(90%)(图4),而其他活动主要是为了维持生计。然而,非农收入是不稳定的,依赖于社会的外生条件,如石油公司对劳动力的需求。相比之下,在Wajosará获得收入的机会更加多样化。约96%的收入来自非农工作、林业和农业,平均每户每周118.6美元。但是,在研究期间,这些活动所产生的收入占总收入的比例极不稳定。如在Zábalo,丛林肉和鱼的销售很少,狩猎和捕鱼是为了满足生存需要(图5)。
丛林肉和鱼类的总收获、家庭内部消耗的丛林肉和鱼类的比例、用于狩猎和捕鱼的劳动时间以及这些活动的成本在Zábalo都大于Wajosará。相比之下,劳动时间专门用来休息Wajosará的农活、野生动物肉和鱼的价格,特别是食品支出更大(表2)。
校准
模型校准使未知参数的数值估计达到观测值(表2)与从理论模型推导出的需求和供给函数预测值之间的一致性。表3显示了用于校准模型的参数值。
弹性
根据使用附录2的方程计算的弹性、经验数据和校准参数,对收获生物量影响最大的外生参数是丛林肉价格、工资和猎物物种的生物学特性,包括生产力和捕获概率(表4)。丛林肉价格每上涨1%,Wajosará和Zábalo的收获生物量分别增加3.1和2.8%。物种生物生产力的同样增长使收获的生物量分别增加了3.4和2.9%,而非农工资的增长使Wajosará和Zábalo的收获生物量分别减少了2.4和1.9%。惩罚和捕猎成本的影响是消极的和无弹性的。这些变量每增加1%,收获的生物量就会以< 1的速率减少。
模拟
将从社区收集到的观测数据纳入校正模型,我们模拟了收获生物量的变化,作为对关键经济参数变化的响应(表5)。对收获生物量影响最大的参数是非农工资。10%的工资增长导致Wajosará和Zábalo的收获生物量分别减少了20%和16%。狩猎成本每增加10%,在各自的群落中收获的生物量就会减少2%和4%。
另外两个感兴趣的外生参数是市场上出售的野生动物肉的价格和被发现出售野生动物肉的概率。假设这两个变量只在猎人决定出售丛林肉时被观察到,模拟分析了对丛林肉销售的影响,而不是对总生物量的收获。价格对出售野味的决定有重要影响。丛林肉价格上涨10%会导致两个社区的肉制品销售量增加约30%(表6)。然而,这一百分比只在Wajosará的绝对数量上相关,那里的实际肉制品销售比例约为11%,这意味着总收获生物量减少了3.7%。相比之下,几乎所有的野味都在Zábalo被消费掉了,所以对野味销售的影响在绝对价值上可以忽略不计。另一方面,野生动物肉价格下降25%使商业狩猎减少了约60%,占Wajosará网站上收获生物量总量的近7%。
鉴于在这些社区中很少有禁止商业狩猎的监管和法规,我们假设了一个基准参数θ= 0.01,即检测出售卖野味的概率设为1%。惩罚K用被没收的野味的市场价值,或者猎人在被捕获时放弃的收入来计算,用卖出的野味的平均重量乘以单位重量的价格来计算。对这一关系的模拟表明,将这一概率提高到2%将使商业狩猎减少44%,而仅提高到5%则意味着商业狩猎实际上将完全停止。
讨论
经济变量对收获生物量的影响
根据该模型,影响土著猎人行为的最重要的经济变量是非农工资。在非法商业狩猎的情况下,丛林肉价格和出售丛林肉被发现的概率对猎人的行为也有重要的影响,但在总收获量方面没有影响。估计的弹性显示出经济理论所预测的迹象。
考虑到厄瓜多尔亚马逊地区目前正在进行的经济发展,工资水平的提高可能会增加土著居民寻找非农工作的动机,从而减少他们花在狩猎上的时间。模拟10%的工资增长会导致社区收获的生物量减少16-20%,而50%的工资增长会使收获的生物量减少> 50%。因此,增加工资将通过减少对野生动物的依赖来改善生物多样性保护,尽管有酬工作的稳定性已被证明对减少野生动物肉消费有更大的影响(Sirén等,2006)。然而,由于更高的工资也增加了狩猎的机会成本,如果猎人使用更有效的方法,减少用于狩猎的时间在理论上的积极影响可能会被抵消,而以前由于成本的原因无法使用这些方法。例如,如果猎人更频繁地使用火器,从狩猎和渔业以外的活动(如农业)中获得的更高收入将对野生动物种群有害(Damania et al. 2005)。我们的数据表明,转向更有效的狩猎技术是可能的,因为在Zábalo和Wajosará, 24%的动物被砍刀或临时矛猎杀。Demmer等人(2002)也报告了经济发展对狩猎普遍性的模糊影响。然而,我们采用的模型并没有捕捉到这种效应。相比之下,Godoy等人(2010)认为增加的收入和野味消费之间不存在显著的联系。根据具体情况,旨在通过提高工资减少狩猎的野生动物保护政策可能会产生意想不到的后果,而且可能不如直接监管有效。
正如预期的那样,狩猎和捕鱼成本的增加减少了生物量的收获量,尽管这种弹性的幅度在所检查的变量中是最低的。鉴于燃料和弹药对猎人和渔民来说是必不可少的,即使这些投入的成本相对大幅增加,也无助于阻止狩猎和捕鱼。模拟成本增加10%导致收获生物量下降< 5%,而增加50%则导致Wajosará和Zábalo分别下降11%和21%。这种影响还可能受到一个事实的影响,即土著社区认为丛林肉是一种必要的商品(Wilkie和Godoy, 2001年)。
与预期一致的是,野味的价格与收获的数量呈正相关。价格上涨50%使Wajosará收获的生物量增加了近25%。相比之下,这种效应在Zábalo几乎为零,在那里,由于社区与市场经济隔绝,丛林肉很少被商业化。较高的价格鼓励出售更多捕获的猎物,从而助长非法贸易,导致对猎物物种的过度开发,并使野生动物种群处于危险之中(Damania et al. 2005)。价格高昂的开放获取系统的不幸后果是自然资源的枯竭和社会福利的恶化(Clark 1990)。相反,模型模拟显示,价格每下降50%,两个社区的野味销量就会减少90%。尽管由于销售比例小,对生物量收获总量的影响有限,但弹性的大小表明,在丛林肉贸易很重要的社区,有效降低丛林肉价格的政策可以促进野生动物保护和可持续生计(Damania et al. 2005)。
鉴于该研究区域几乎不存在对狩猎的监测和监管,我们进行了进一步的模拟,以评估改进的监测的效果,发现商业狩猎在超过5%的探测概率阈值后几乎消失。这些结果表明,监测的作用比Damania等人(2005年)报告的更为有效,并表明一个健全的检查制度将对非法丛林肉贸易产生重大影响。这种制度需要了解进入市场的模式和机制,理想情况下应包括某种形式的以社区为基础的控制。就野生动物保护而言,与降价一样,监管的好处将是有限的,因为目前出售的野生动物肉的比例很小,但如果社区与市场的联系越来越紧密,这是未来的一个重要工具。
不同社区之间野生动物收割的差异
两个社区在市场整合和相对于保护区的位置方面的差异有助于解释丛林肉收获和捕鱼水平的不同。结果表明,野生动物来源的食物是研究地区大多数土著家庭的主要蛋白质来源。狩猎提供了平均0.10和0.33公斤的人−1一天−1分别在Wajosará和Zábalo上发布了大量的野生动物肉。后者与体重0.44公斤的人相似−1一天−1厄瓜多尔的一个阿伊Cofán社区报告了未经屠宰的丛林肉的收获情况(谢尔,1997年)。相比之下,在Wajosará观察到的量远远低于0.42公斤的人−1一天−1一个独立的Secoya土著社区报告了大量被屠宰的丛林肉(Vickers 1980)。在我们的研究和早期的研究之间,Secoya社区的丛林肉消费的差异可以解释为Wajosará地区的森林覆盖和周围景观的下降,这是由于农业前沿的发展(主要是油棕种植园),以及这些社区比过去更大程度地融入商品和劳动力市场(Vickers 1991)。相比之下,Zábalo在地理和经济上相对孤立,仍然高度依赖蛋白质的自然供应。
捕鱼似乎没有狩猎重要,而且明显是季节性的。这种活动在9月和一年的头几个月的旱季更常见,这时一些物种大量游向阿瓜里科河,在较小的支流产卵(E. de la Montaña,个人观察).这种可靠的鱼类资源在旱季的每月捕捞数据中得到了反映,那时捕捞的鱼类生物量与通过狩猎捕获的生物量相似,甚至更高。两种蛋白质来源之间的互补性在Zábalo比在Wajosará更明显,这同样是因为前者社区对自然资源的依赖程度更高。
对进一步研究的见解
基础模型的进一步迭代改变了家庭同质性、具有代表性的游戏物种和单一狩猎技术的假设,这是一个有前途的研究领域。尽管两个社区的市场整合和森林保护水平不同,但结果趋于一致,这支持了该模型可以应用于严重依赖狩猎和捕鱼的相似群体的想法。根据特定的社会经济和狩猎环境,可以修改模型约束,以纳入不同的生产活动、劳动条件和野生动物捕获技术。我们建议根据保护状况(即非受威胁或濒危物种)和社会经济因素(如市场价格和消费者对丛林肉的偏好)来选择要分析的物种。通过调整生产函数,可以更好地了解特定野味物种的生物学特性的影响。该模型还允许通过包含特定的生产功能来合并不同类型的狩猎技术。尽管增加物种或技术的数量将大大增加模型的复杂性,并需要更多的经验信息,但该模型为洞察猎人的经济决策提供了一个强大的工具。
该模型的一个局限性是忽略了农业活动。对大多数社区来说,自给农业是日常活动的一部分,需要家庭成员投入时间。考虑到与收集太多数据和建模几个决策变量相关的复杂性,我们决定将重点放在钓鱼和狩猎上,因为它们的行为非常接近。此外,以前对该研究领域的观察表明,用于农业的时间往往是固定的和非弹性的,相对于其他活动,如打猎或捕鱼。尽管忽略了农业活动,但该模型的关键含义并没有受到很大影响。在用于农业活动的时间受狩猎劳动需求影响的情况下,可以调整模型以将其包括在内。
结论
制定一个土著家庭经济的简单模型的目标是为分析和制定政策建立坚实的理论基础,以促进可持续的丛林肉提取和当地粮食安全。据我们所知,这是拉丁美洲第一个模拟保护和发展政策对猎人行为影响的研究,并讨论了对生物多样性保护和土著生计的影响。19个月的数据收集使我们能够捕捉到影响生物系统和相关经济活动的季节变化。结果表明,在研究的社区中,非农工资的变化对收获的生物量影响最大。在与市场经济结合更为紧密的社区,野生动物肉价格和对非法贸易的惩罚对收获的生物量的影响比孤立的社区更大。研究结果与同类研究结果一致,可在其他地区推广应用。然而,由于影响野味消费的因素是复杂的,而且是在非常地方的层面上运作的,我们敦促在对结果进行广泛的概括和讨论管理影响时要谨慎。虽然这里给出的结果是基于对现实的简化,但模型约束可以调整以反映更复杂的社会经济和狩猎背景。该模型的开发将使研究人员能够进一步深入了解猎人的经济决策,并更充分地了解影响类似社区丛林肉消费的因素。
致谢
厄瓜多尔高等教育、科学、技术和创新秘书处(SENESCYT)的Prometeo项目、西班牙国际发展合作署(MAEC-AECID)、美国国际开发署(USAID)资助的安第斯亚马逊保护倡议(ICAA)和保护战略基金(CSF)提供了财政支持。国际林业研究中心(CIFOR)为这篇文章的出版提供了资金支持。我们衷心感谢Wajosará和Zábalo的人们,特别是那些作为现场助理合作的人。我们也感谢利兹·奥尔蒂斯和雪莉·祖利塔对数据分析的支持。两位匿名审稿人提供了宝贵的意见,提高了稿件的质量。
文献引用
R. E.博德默,E. P.洛扎诺,方T. G. 2004。秘鲁亚马逊地区野生动物利用的经济分析。191 - 207页在K. M.西尔维乌斯,R. E.博德默和J. M. V.弗拉戈索,编辑。人与自然:南美洲和中美洲的野生动物保护。哥伦比亚大学出版社,美国纽约。
布拉舍尔斯,j.s, c.d.戈尔登,K. Z.温鲍姆,C. B.巴雷特,G. V.奥凯洛,2011。非洲农村野生动物消费的经济和地理驱动因素。美国国家科学院院刊108(34): 13931 - 13936。http://dx.doi.org/10.1073/pnas.1011526108
布鲁克斯,t.m., R. A. Mittermeier, G. A. B. da Fonseca, J. Gerlach, M. Hoffmann, J. F. Lamoreux, C. G. Mittermeier, J. D. Pilgrim, A. S. L. Rodrigues. 2006。全球生物多样性保护优先事项。科学313:58 - 61。http://dx.doi.org/10.1126/science.1127609
布朗,d . 2003。野味与扶贫:对发展政策的影响。ODI野生动物政策简报海外发展研究所,伦敦,英国。(在线)网址:http://www.odi.org/publications/2427-bushmeat-poverty-alleviation-implications-development-policy
克拉克,1990年。数学生物经济学:可再生资源的最佳管理。Wiley-Interscience,纽约,美国纽约。
Damania, R., E. J. Milner-Gulland和D. J. Crookes. 2005。丛林肉狩猎的生物经济学分析。英国皇家学会学报B272:259 - 266。http://dx.doi.org/10.1098/rspb.2004.2945
de la Montaña, E. 2013。Cacería厄瓜多尔人的生存环境indígenas de la Amazonía。Ecosistemas22(2): 84 - 96。http://dx.doi.org/10.7818/ecos.2013.22-2.13
de Merode, E. K. Homewood和G. Cowlishaw. 2004。丛林肉和其他野生食物对刚果民主共和国生活在极端贫困中的农村家庭的价值。生物保护118(5): 573 - 581。http://dx.doi.org/10.1016/j.biocon.2003.10.005
Demmer, J., R. Godoy, D. Wilkie, H. Overman, M. Taimur, K. Fernando, R. Gupta, K. McSweeney, N. Brokaw, S. Sriram, T. Price. 2002。收入水平能否解释游戏丰富度的差异?在洪都拉斯的两个村庄进行的实证检验。生物多样性和保护11(10): 1845 - 1868。http://dx.doi.org/10.1023/A:1020305903156
法J. E.和C. A.佩雷斯。2001。非洲和新热带森林狩猎脊椎动物的提取:洲际比较。203 - 241页在j·d·雷诺兹,m·梅斯,k·h·雷德福,j·g·罗宾逊,编辑。保护已开发的物种。剑桥大学出版社,英国剑桥。
法,j.e, C. A.佩雷斯,J. Meeuwig. 2002。热带森林中的丛林肉开发:洲际比较。保护生物学16(1): 232 - 237。http://dx.doi.org/10.1046/j.1523-1739.2002.00275.x
弗兰岑,m . 2006。评估狩猎的可持续性:三个华欧拉尼社区的收获情况比较。环境保护33(1): 36-45。http://dx.doi.org/10.1017/S0376892906002712
戈多伊,E. A. Undurraga, D. Wilkie, V. Reyes-García, T. Huanca, W. R. Leonard, T. McDade, S. Tanner, V. Vadez,和TAPS玻利维亚研究团队。2010。玻利维亚土著亚马逊人的财富和实际收入对野生动物消费的影响:基于纵向家庭数据的年度趋势估计(2002-2006)。动物保护13(3): 265 - 274。http://dx.doi.org/10.1111/j.1469-1795.2009.00330.x
国家研究所Estadística y Censos, 2010。Censo de población y de vivienda:结果。国家研究所Estadística y Censos,厄瓜多尔基多。(在线)网址:http://www.inec.gob.ec/cpv/
Iwamura, T, E. F. Lambin, K. M. Silvius, J. B. Luzar和J. M. V. Fragoso. 2014。土著土地上狩猎和自给农业的基于主体的建模:理解社会和生态系统之间的相互作用。环境建模及软件58:109 - 127。http://dx.doi.org/10.1016/j.envsoft.2014.03.008
李维,T., G. H. Shepard Jr., J. Ohl-Schacherer, C. A. Peres和D. W. Yu. 2009。模拟秘鲁马努国家公园土著狩猎的长期可持续性:对亚马逊地区景观规模管理的影响。应用生态学杂志46(4): 804 - 814。http://dx.doi.org/10.1111/j.1365-2664.2009.01661.x
Milner-Gulland, E. J, E. L. Bennett和保护生物学学会2002年年会野生肉类小组。2003。野味:大局。生态学与进化趋势“,18(7): 351 - 357。http://dx.doi.org/10.1016/s0169 - 5347 (03) 00123 - x
迈尔斯,R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, J. Kent, 2000。生物多样性热点是保护重点。自然403:853 - 858。http://dx.doi.org/10.1038/35002501
奥尔森,D. M.和E.迪纳斯坦。2002。全球200强:全球保护的优先生态区域。密苏里植物园年鉴89(2): 199 - 224。http://dx.doi.org/10.2307/3298564
佩雷斯,2000年。评估亚马孙雨林多个地区自给狩猎的影响和可持续性。页面31-56在j·g·罗宾逊和e·l·班尼特,编辑。寻找热带森林的可持续性。哥伦比亚大学出版社,美国纽约。
Rivadeneira, c . 2007。producción库亚贝诺动物保护区。页面形成反差在ECOLAP和MAE,编辑。Guía del patrimonio de áreas厄瓜多尔自然保护。ecfund, FAN, DarwinNet和IGM,厄瓜多尔基多。
罗宾逊,J. G.和K. H.雷德福,1991。新热带森林哺乳动物的可持续收获。415 - 429页在j·g·罗宾逊和k·h·雷德福,编辑。新热带野生动物的利用和保护。芝加哥大学出版社,美国伊利诺斯州芝加哥。
Schel l . 1997。Cacería厄瓜多尔的脊椎动物和陆地的建议和建议,在组织的国家和地区,conservación de物种和社会Cofán de Zábalo, Producción Faunística de Cuyabeno保留地,Amazonía厄瓜多尔。论文。厄瓜多尔,基多,Pontificia大学Católica。
谢泼德,G. H. Jr, T. Levi, E. G. Neves, C. A. Peres, D. W. Yu. 2012。古代和现代亚马逊地区的狩猎:对可持续性的重新思考。美国人类学家114(4): 652 - 667。http://dx.doi.org/10.1111/j.1548-1433.2012.01514.x
塞拉,R。编辑器。1999.Propuesta初级的联合国系统clasificación de la vegetación厄瓜多尔大陆。INEFAN-EcoCiencia,基多、厄瓜多尔。(在线)网址:http://www.ecociencia.org/archivos/RSierra_PropVegEcuador_1999-120103.pdf
A. H. Sirén, J. C. Cardenas和J. D. Machoa. 2006。热带森林收入与狩猎之间的关系:实地经济实验。生态和社会11(1): 44。(在线)网址://www.dpl-cld.com/vol11/iss1/art44/
Sirén, A. P. Hambäck, J. Machoa. 2004。在评价狩猎时包括空间异质性和动物分散:一个亚马逊社区的模型分析和实证评估。保护生物学18(5): 1315 - 1329。http://dx.doi.org/10.1111/j.1523-1739.2004.00024.x
维克斯,1980年。亚马逊狩猎产量与定居年龄的函数分析。页面7-30在R. B. Hames和K. M. Kensinger,编辑。新热带地区的狩猎和捕鱼研究2。本宁顿学院,美国佛蒙特州本宁顿。
维克斯,1991。在亚马逊印第安人领地的十年狩猎产量和猎物组成。53 - 81页在j·g·罗宾逊和k·h·雷德福,编辑。新热带野生动物的利用和保护。芝加哥大学出版社,美国伊利诺斯州芝加哥。
Vié, J. C.希尔顿-泰勒和S. N.斯图尔特编辑。2009.野生动物在一个变化的世界:2008年IUCN濒危物种红色名录的分析。世界自然保护联盟、腺、瑞士。(在线)网址:https://portals.iucn.org/library/efiles/documents/RL-2009-001.pdf
威尔基,D. S.和R. A.戈多伊,2001。低地美洲印第安人社会丛林肉需求的收入和价格弹性。保护生物学15(3): 761 - 769。http://dx.doi.org/10.1046/j.1523-1739.2001.015003761.x
C. Urgilés, E. Suárez。2009.厄瓜多尔亚马逊的舒阿尔人狩猎哺乳动物:这是可持续的吗?大羚羊43(3): 375 - 385。http://dx.doi.org/10.1017/s0030605309001914
