

去pdf本文的版本
莫拉,F., P. Balvanera, E. Garc—a-Frapolli, A. Castillo, J. M. Trilleras, D. Cohen-Salgado,和O. Salmer—n。2016.热带干燥林区生态系统服务与可持续性替代途径之间的权衡。生态和社会(4): 45。
https://doi.org/10.5751/ES-08691-210445
热带干燥林区生态系统服务与可持续性替代途径之间的权衡
摘要
设计旨在可持续资源管理的战略需要理解所涉生态系统服务之间的权衡,以确定驾驭它们的适当方法。我们评估了墨西哥热带干燥森林中用于牧牛的牧草生产与维持碳储量或树木多样性之间的权衡。通过确定其在场地和景观尺度上的效率边界,评估了对服务之间的权衡。我们还评估了在当前和假设土地管理条件下的服务结果。我们发现,在站点尺度上,饲料和碳储量之间以及饲料和树种丰富度之间存在明显的权衡。在景观尺度上,效率边界呈凹形,而在饲料-物种丰富度情况下,效率边界的权衡不明显。我们对当前服务供应水平的估计显示,相对于沿效率边界的最大可行值,C存量减少了18-21%,饲料生物量减少了41-43%。降低这种低效率的最佳管理策略的选择取决于森林砍伐程度:当森林砍伐程度较低时次生林更新最适合,而当森林砍伐程度较高时提高牧场的饲料生产力最适合。考虑到有利生态因素的范围,以及考虑到当地利益攸关方的要求和偏好,用草料树和次生林进行牧草丰产是实现可持续性和平衡生态和社会可持续性的潜在管理选择。考虑到在整个热带地区都发现了类似的权衡,这项工作有助于协调热带森林的维护及其对可持续农村生计的利用。介绍
人类常常以减少调节或支持服务为代价来增强供应服务的交付(Rodríguez等人,2006年,Raudsepp-Hearne等人,2010年)。在热带干林(TDF)地区,土地所有者经常砍伐他们的森林并为养牛建立牧场(Burgos和Maass 2004, Sánchez-Azofeifa和Portillo-Quintero 2011),因为牲畜构成了一种私人商品,可以在成熟的市场上交易,从那里可以获得短期净回报,但也因为牲畜的功能很像一个“储蓄账户”,可以用于获得现金以应对金融紧急情况(Gerritsen等人,2007年,Casas等人,2008年)。然而,森林砍伐导致温室气体排放(Achard et al. 2014)、地方和区域气候调节失衡(Anderson-Teixeira et al. 2012)、生物多样性和相关服务的丧失(de Albuquerque et al. 2005, Soto 2009)和土壤生产力下降(Maass et al. 1988, Trilleras et al. 2015)。所有这些影响都与管制或辅助服务的丧失有关,其中大多数是公共产品(McKean 2000),土地所有者得不到经济补偿或有较长的归还时间。
在不影响监管或支持服务(包括生物多样性)的情况下,对改造后的景观进行持续服务供应的管理仍然是一项挑战(Polasky等,2008,Tallis等,2008,Brown和Mumby, 2014, Gilroy等,2014)。识别服务之间的权衡和协同作用对实现这一目标至关重要(Raudsepp-Hearne等人2010,Ruijs等人2013)。例如,碳储量和生物多样性的维持是热带干燥森林景观提供的两项最重要的服务(Balvanera等人,2011年,portilo - quintero等人,2015年),这取决于推动森林砍伐的地方尺度上的权衡与调节森林砍伐的景观或区域尺度上的社会经济因素之间的相互作用。在当地范围内,以牛为饲料的外来草的生长与森林下层植被中减少的阳光不相适应(Beale 1973, sirceli和Naeem 2012)。因此,森林被清除以增加草的产量。然而,考虑到牧牛带来的低收入、劳动力供应的限制以及农村家庭对多种经济活动的依赖(Lambin et al. 2003),放牧并不是整个景观的主导。此外,小土地所有者重视保护其森林的某些部分,以获得饲料和森林产品的替代来源(Harvey和Haber 1999, Barrance等人,2003)和森林对维持生命的贡献(Castillo等人,2005)。因此,查明和量化这种权衡对推动旨在平衡地方私人利益和区域或全球公共利益的管理战略的设计至关重要。
Cavender-Bares等人(2015)最近开发了一个用于评估生态系统服务之间权衡的概念框架。两种服务之间的权衡可以用效率边界来描述,效率边界是在目前的生物物理和管理条件下,两种服务可以获得的最大可行值的组合。可以评估与此最佳条件相关的系统当前状况,即当前效率,从而识别与特定管理实践相关的路径,以及使系统能够或阻止系统向这一边界移动或沿着这一边界移动的生态和社会因素(King et al. 2015)。
我们在这项研究中应用了该框架来评估墨西哥西部热带干燥森林地区的服务之间的权衡。我们使用了来自一个长期社会生态研究站点(Castillo等人2005,Maass等人2005,Martínez-Ramos等人2012)的数据和模拟方法,在站点和景观尺度上评估牛的生产、碳储量维持和生物多样性保护之间的权衡。我们提出了以下具体问题:(1)在场地尺度上,养牛生产、碳储量维持和生物多样性保护之间的权衡是什么性质的?(2)在比较场地和景观尺度时,这些权衡如何变化?(3)当前的景观配置对于提供这些服务的效率如何?最后,我们讨论了可替代的管理条件和具有更高可持续性潜力的相关途径,其基础是对促进或抑制每种途径的生态和社会因素的讨论。
方法
研究区域
这项研究集中在墨西哥太平洋海岸的Chamela-Cuixmala生物圈保护区(CCBR)周围地区(19°23 ' 02″-19°36 ' 51″N, 104°56 ' 07″-105°03 ' 36″W;图1).该地区年平均气温为24.6℃,年平均降水量为788毫米。降水强烈集中在11月至5月之间(García-Oliva et al. 2002)。主要地形是丘陵和平原,分别占景观的89%和11% (Castillo et al. 2005)。丘陵有中等到极陡的斜坡和营养不良的土壤(Cotler等,2002年),热带干燥森林生长(Durán等,2002年)。平原包括沿着主要河流和季节性溪流的泛滥平原和山谷,土壤更深厚,营养丰富,热带半常青林生长在土壤上(Durán et al. 2002)。
尽管自1993年以来CCBR一直致力于严格的保护和科学研究目的,但在过去50年里,邻近地区因农业活动而发生了不同程度的变化(Maass et al. 2005)。农民们砍伐和烧毁了部分土地,在平原上种植作物,并在山上建立牧场放牧(Burgos和Maass 2004年,Cohen Salgado 2014年)。然而,牧场有时被废弃,然后被次生林覆盖,次生林可以与残存的原生林一起用作木材、木桩和柴火提取,以及养牛(Rendón-Carmona et al. 2009, Cohen Salgado 2014)。
生态系统服务评估
我们收集了位于CCBR内部和周围的55个站点的数据集,对这些站点的饲料可用性(在这里用作牛产量的代表)、地上碳储量和树种丰富度(用作生物多样性的代表)进行了估计(Martínez-Harms 2010, Quijas 2012, Mora等人2015,Trilleras等人2015)。这些地点都以三种不同的土地覆盖类型之一为特征:活性牧场(28)、次生林(14)和原生林(13),所有这些都是在TDF历史存在的生物物理条件下出现的。牧场的管理制度从研究前1-3年才建立的、没有后期维护的牧场,到30年前建立的、遭受过多次刀耕马种事件的牧场(Trilleras et al. 2015)。次生林分布范围从冠层较短(≈2 m)到结构和组成与原生林相似的森林。原生林包括采伐林产品或放牧的地点,以及保护区内受到严格保护的地点(Martínez-Harms 2010, Quijas 2012, Mora et al. 2015)。
饲料可用性以1 m²地块内胸径小于1 cm的所有植株的生物量近似计算,每个地块2、3和5个样本(分别为19、12和24个地块,根据数据源而定)的平均值。在牧场,这种生物量主要由外来草类组成,而在原生林和次生林中,它由大量的本地草本植物、小灌木和典型的森林下层树苗组成(Maza-Villalobos等,2011;M. R. Ocampo-Dominguez 2012,未出版的手稿).这个定义包括被牛最积极消耗的植物层;由于缺乏数据,它不包括牛在距离地面1米以上或掉落时吃掉的叶子和果实(特别是豆荚)。样品经烤箱干燥以确定干质量。我们报告了每个站点的平均生物量。
地上碳储量(以下简称C储量)由树木(所有活灌木和胸径≥1 cm的乔木)和饲料组分估算。对于树木组成部分,树木普查使用两种地块大小:400平方米的方形地块(19个站点)和1000平方米的矩形地块(36个站点)。对于第一个地块大小,> 1.0,> 2.5,> 0 cm的个体。胸围高度的直径分别在25、100和400 m²的嵌套图中测量。对于第二个地块大小,在500、1000和1800 m²的嵌套矩形地块中测量个体> 1.0、> 2.5和> 10.0 cm DBH。使用树木生物量与胸径的异速生长方程估算地上活树生物量(Martínez-Yrízar等,1992年)。饲料生物量的估计如上所述。根据相应的碳浓度值,将树木和饲料组分的生物量转化为C树(Jaramillo et al. 2003)。
利用同一树种普查估算树种丰富度。为了使我们对不同地点的物种数量的估计具有可比性,我们使用了一种称为稀疏的标准化方法(Gotelli and Colwell 2001),该方法考虑了不同地点之间树木密度和采样协议的差异。对于普查中有超过30个个体的2.5 cm胸径的站点(27个站点),使用纯素库对R进行稀疏处理(Oksanen et al. 2011)。对于个体数小于30个的站点(28个站点,均为当前牧场,个体数小于10个),采用实测树种丰富度。
权衡的评估
我们定义了场地和景观尺度的效率边界,以评估饲料有效性和地上碳储量或树种丰富度之间的权衡(Cavender-Bares et al. 2015)。
Site-scale权衡
在站点尺度上,我们利用55个采样站点的数据,将碳储量-饲料和树木多样性-饲料效率前沿定义为其二元数据簇的上限。我们使用了一种简单的启发式方法,包括拟合一个四参数逻辑函数到数据集群,这样没有数据点位于函数形成的曲线之上:
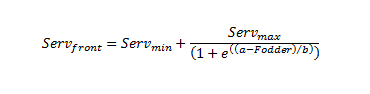 |
(1) |
该函数将效率边界定义为s型衰减形式。服务公司前面边界上的C资源或物种丰富度值是的函数吗饲料.的总和服务公司最小值而且服务公司马克斯的渐近初值的估计值服务公司前面当饲料接近最小值时。参数b用参数定义衰减速率一个定义拐点发生时的素材值。最后,服务公司最小值的渐近最小值服务公司前面当饲料增加到最大值时。
选择最适当的参数估计通过重复拟合遵循数值优化程序。我们首先拟合10的函数4参数组合(每个参数有10个初始值)使用“nls2”包和R (R Development Core Team 2015)中的“蛮力”方法。然后我们选择残差都为负的拟合,即所有的点都位于直线以下。最后,我们选择残差之和大于99的拟合th百分位,即最小化点与线之间的总体距离。这些拟合参数的结果值范围用于定义下一轮拟合的后续初始值。重复这个过程,直到后续模型的残差之和的差异小于1%。
对每个数据集群运行两种不同的拟合。第一个拟合包括所有数据点(完全数据拟合),而第二个拟合排除了C种群和物种丰富度值中最极端的5%,以消除极值的影响(95%数据拟合)。为了识别这些极端数据,我们将饲料范围细分为三个百分位类别(0-33%、36-66%和67-100%),并在每个类别中排除了最高的C储量和物种丰富度值。
景观尺度的权衡
景观被定义为约477平方公里的区域,其中包括CCBR(约132平方公里)和5公里缓冲区内的相邻区域(约345平方公里),55个站点中的大多数位于该区域。这个缓冲面积是通过假设备用多边形为20.0 x 6.6 km的完美矩形来计算的。景观被细分为0.24 ha样地,因为这是计算物种丰富度数据的单位。
通过模拟逐级森林砍伐过程,在景观尺度上评估了效率边界的形状。该模拟只包括两种土地覆盖类型:原生林和牧场。尽管我们承认次生林是该地区重要的土地覆被类型,但由于在这方面的物种发生率数据不足,次生林没有被纳入效率前沿的替代选择。我们还假设景观只包括丘陵地区,那里饲料生产集中(Maass et al. 2005)。
在开始模拟之前,我们为每个地块分配了原始森林和牧场覆盖下的饲料和C存量值。由于我们试图定义效率边界,因此从55个采样点中的52个(95%数据曲线以上的点被排除在外)中观察到的最高饲料和C存量值所定义的正态分布中随机分配值。最高的值定义为每个服务和覆盖类型组合的第三四分位数以上的值。我们使用它们的均值和标准差值来定义正态分布(表1)。我们检查了分配的值都在95%数据边界以下。当情况并非如此时,我们使用边界函数对值进行强制,以估计其指定饲料水平对应的C库存量。
物种组成不能按照相同的标准进行分配,因为每种覆盖类型的站点数量有限,无法充分估计物种出现的可能性。相反,对于原生林,我们使用了21个0.24公顷的原生林地块的物种组成的公开数据,包含144种树种(Durán et al. 2006),并计算了一个地块内每种物种的存在概率。然后,我们使用这些概率和随机数生成器来定义一个图中物种的存在/缺失:每当随机数(在0到1之间)等于或大于该物种存在的实际概率时,它就被指定为存在,否则就被认为不存在。对所有144个物种和整个景观的所有地块重复了这个过程。对于牧场,我们自己的数据(样本量小)和其他可用数据(Durán et al. 2006)都不足以评估物种组成。因此,我们没有在景观物种丰富度的量化中包括牧场。
模拟阶梯式森林砍伐过程始于完全被原始森林覆盖的景观。地块被随机分配在CCBR内外的位置。在模拟过程中,首先砍伐保护区以外的地块。只有在保护区外的景观完全转变为牧场后,保护区内的地块才被认为符合砍伐条件。根据从原生林转变为牧场时,每单位碳损失的饲料收获比,选择有砍伐潜力的地块。在每个模拟步骤中,砍伐占比最高的10%的剩余原生林样地。当所有的景观都转换为牧场时,模拟结束。这个过程使我们能够找到C库存饲料案例的确切边界。然而,对于种属饲料的例子,该程序只产生了精确边界的近似值。
在每个模拟步骤结束后,我们计算了景观尺度的饲料、C种群和物种丰富度值。通过简单地将所有地块的饲料和C存量值相加(作为地块土地覆盖的函数),计算出它们的地块级别值。景观层面的物种丰富度定义为在整个景观中至少1/21的样地(9455个样地)中存在的物种数量,这对应于Durán等人(2006)提出的最小物种存在概率。这种可能性意味着,考虑到该地区的树木密度和花卉生产的最小树木尺寸,研究区域内的种群规模约为10,000个可繁殖个体(Bullock 2002年)一个,b2004年,杜兰)。我们认为这样的成虫密度足以维持任何树种的存活种群。
最后,通过绘制景观C种群和物种丰富度随饲料变化的函数值,确定景观效率前沿。模拟过程重复1000次。由于不同绘图的结果非常相似,所以只显示平均值。所有的分析都是使用R软件(R Development Core Team 2015)进行的。
目前园林饲料供应和C库
我们根据CCBR以外的两个对当前森林砍伐的独立估计,估算了当前景观级别的饲料供应和C库存维持情况。根据卫星图像的解释,整个景观的“低森林砍伐”估计为17% (Sánchez-Azofeifa等,2009年)。根据土地所有者对森林砍伐的报告看法,估计有61%的景观被砍伐(Sánchez-Azofeifa et al. 2009, Cohen Salgado 2014)。
在每一个森林砍伐场景中,地块被随机分配为森林或牧场覆盖。森林样地根据其对总森林覆盖的贡献比例被划分为原生林或次生林。我们依靠的是次生林覆盖率的粗略估计,该估计来自30个不同的土地所有者对其财产上存在的不同类型的覆盖的感知(F. Mora,未发表的数据).据估计,总共有36%的森林是次级森林,即以前为农业目的而砍伐后被遗弃的地方。CCBR内的地块总是分配给原始森林覆盖。根据土地覆盖定义,我们为每个地块分配其当前饲料和C库存值,使用的程序类似于前面描述的定义景观级边界的程序。所用饲料和C料的平均数和标准差估计数载于表1。这些参数估计是根据我们所有的数据计算出来的,使用CCBR以外的站点;它们被认为是该区域目前森林和牧场管理状况的代表(用于牧牛和提取林产品的森林)。我们通过汇总整个景观中每种服务的对应地块级别值,计算了当前两种森林砍伐估算的景观级别饲料供应和c库存维持。
最后,我们通过模拟三种假设但可行、社会满意的管理场景(总是假设土地覆盖没有进一步变化),评估了不同的管理活动如何推动景观服务供应向效率前沿发展:
- 允许次生林再生:次生林通过允许再生达到其最大的C林和饲料值(表1),而在当前条件下继续使用原生林和牧场。
- 通过持续的现行做法进行牧场管理:通过现行土地所有者的做法来管理牧场,众所周知,这些做法可以提高饲料生产力,例如反复的刀耕火种事件,因此在持续管理30年后,观察到的饲料价值达到最大值(表1)(Trilleras等人,2015年)。
- 次生林再生和通过现行做法进行牧场管理:这是前两种管理条件的结合。
我们没有在景观尺度上评估当前的物种丰富度,因为数据在充分估计原生林(未受保护)和次生林遗址下的物种出现概率方面存在局限性,特别是考虑到热带树木之间的演替生态位的存在,以及该TDF中物种组成沿演替的快速变化(Letcher et al. 2015, Mora et al. 2015)。
结果
Plot-level权衡
我们发现,在站点水平上,饲料和C种群之间,以及饲料和物种丰富度之间存在很强的权衡。排除5%最极端的数据明显改变了前沿的位置,尤其是C种群,尽管它没有从质的上改变它们的形状(图2)。在饲料值约2 Mg ha时,C种群和丰富度都出现了逐步下降-1对于95%的数据拟合。然后,效率边界稳定在5.6 Mg C ha-1(表2)。可达到的最大C库存量(服务公司最小值+服务公司马克斯)从42.3 Mg ha降至33.4 Mg ha-1剔除极端数据后,丰富度从23.0下降到19.3种(图2b,表2)。
原生林C储量最高,饲料储量最低,而牧场则相反(表1)。与原生林相比,次生林C储量减少,而饲料可用性略有增加(图2a,表1)。在物种丰富度方面也发现了同样的一般模式,尽管次生林的物种丰富度与原生林相似甚至更高(图2b)。C种群与丰富度高度相关(Spearman rho = 0.87,年代= 3710, p < 0.001;附录1)。
景观水平的权衡
景观层面的权衡要比地块尺度上的权衡明显得多。饲料和C群的景观层效率边界呈微凹状(图3a)。沿效率边界,当景观完全被原生林覆盖时,C储量最大值为1.47 Tg,饲料价值最小值为41.8 Gg(图3a中点“a”)。储量占最大C储量(0.56 Tg C;图3b中的“d”点)。在完全毁林条件下,只有14.2% (0.21 Tg C)的C被保存,达到了饲料的最大值382 Gg(图3a中的“e”点)。
饲料和木本物种丰富度效率边界严格凹(图3b)。整个景观沿边界估计的最大物种数量为135个,在完全森林覆盖下发现(图3b中的“a”点)。生物圈保护区在保护这种多样性方面发挥着重要作用,因为如果假设效率最高,仅生物圈保护区就平均保护了87种物种(最大值的64%)(图3b中的“d”点)。在完全砍伐森林的情况下没有发现任何物种,因为我们在景观尺度的物种丰富度量化中没有包括牧场(图3b中的“e”点)。
景观水平的当前饲料和C库存
在效率边界上,两种目前的森林砍伐估计与景观尺度上的C储量水平相关联。根据目前森林砍伐的低估值和高估值,分别损失了10.2%至37.4%的碳储量(图3a,点“b”和“C”)。相比之下,在森林砍伐估计值之间没有观察到边界物种丰富度的差异;在这两种情况下,观察到的物种丰富度最大值仅减少了14个物种(10.4%)(图3b,点“b”和“c”)。
目前景观水平的饲料有效性和C库被发现远未达到最佳状态。在森林砍伐程度较低的情况下,估计整个景观可提供57克饲料和1.04克碳储量(图3a,点“b”)咕咕叫),分别对应沿边界(图3a,点b)。在高度砍伐的情况下,估算出饲料的121 Gg和0.75 Tg C(图3a,点“C”咕咕叫),分别对应效率前沿值的57.0%和82.0%(图3a,点“c”)。
尽管每种管理方案的相对效果取决于森林砍伐状况,但评估的所有三种管理方案都与将当前服务供应转向景观规模效率前沿有关。次生林恢复与C储量的增加有关,低森林砍伐(0.09 Tg C)大于高森林砍伐(0.04 Tg C;图3a),而只引起饲料值的微小变化(< 1 Gg)。相比之下,在低森林砍伐和高森林砍伐估计下,提高牧场效率使饲料供应量分别增加了31.2和87.0 Gg。在这种情况下,碳储量也增加了,在低森林砍伐和高森林砍伐估计下分别增加了0.01和0.04 Tg C。因此,在次生林再生和提高牧场效率的情况下,对于高砍伐估计的C储量的增加是相同的(图3a)。这两种管理战略的结合大大改善了饲料供应和碳储量,特别是在森林砍伐估计较高的情况下,与森林砍伐估计较低的情况相比,到边界的距离较短(图3a)。
讨论
两个空间尺度的效率前沿
场地尺度与景观尺度之间的效率边界形式的显著差异表明,在不同的空间尺度上,服务供给存在着不同的过程。在地块尺度上发现的凸效率边界很可能反映了光作为植物生长的限制资源的作用。目前,森林砍伐是增加引进的、需要光照的草的饲料供应的必要条件,因为在低饲料水平下,C类植物储量和物种丰富度急剧下降表明了这一点。此外,如果允许灌木和树木在牧场上生长,林冠覆盖在短短几年内就会迅速恢复(Lebrija-Trejos et al. 2008, Mora et al. 2015)。冠层覆盖度的增加减少了下层植被的光照可用性(Lebrija-Trejos et al. 2011, Pineda-García et al. 2013),从而排除了这些草。农民们清楚地意识到这种权衡,并投入时间、精力和金钱定期砍伐和烧毁他们的牧场,以避免木本物种的生长(Burgos和Maass 2004年,Trilleras等人2015年)。因为物种丰富度与个体密度部分相关(Gotelli和Colwell 2001),与森林砍伐相关的树木密度的大幅减少也意味着树木物种丰富度的急剧下降。
缩放到场景时,效率边界形状的变化来自于跨站点聚合服务的方式和这些站点之间服务供应冗余水平的差异。碳储存不是一项多余的服务,即,在一些地块通过砍伐森林而损失的碳储存并不能通过保持其他地块的森林来补偿。因此,C种群的效率边界反映了采伐地块的选择方式:典型的凹曲线产生于首先采伐那些牧场生产力最高、碳损失最低的地块。一个类似的过程被用于定义其他效率前沿曲线(Polasky et al. 2008, Cavender-Bares et al. 2015)。事实上,曲线只有轻微的凹,这是由于不同地块的饲料获得与碳损失的比率相对相似。物种丰富度边界的凹陷更为明显,因为这是一种多余的服务,即在整个森林砍伐过程中,残余森林缓冲了物种的损失。只有当森林的重要部分已经消失时,存在物种的地块的比例才会低于规定的灭绝阈值。其他陆地和水生环境也描述了这种模式(Polasky et al. 2008, Brown and Mumby 2014)。
我们的数据证实了众所周知的森林保护(占碳储量和物种丰富度的大部分)与依赖森林砍伐和畜牧生产的农民生计之间的权衡(Cohen Salgado 2014, Salmerón 2015)。然而,养牛场在该地区的经济效益非常低,甚至会给农民造成经济损失(Salmerón 2015)。牲畜被视为一个“储蓄账户”,这一基本原理可能会维持牛牧场的持续经营,在金融紧急情况下(农发基金2004年,Gerritsen et al. 2007年,Casas et al. 2008年),或在数十年政府促进畜牧生产的项目之后,由于某种历史“惯性”(Maass et al. 2005年,Borrego and Skutsch 2014年),以及与社会声望或对“牧场主”的尊重相关的文化原因(Shadow 2002年,Gerritsen et al. 2007年)。结果是,随着森林砍伐的加剧,碳储存的公共利益已经丧失,有时甚至没有从畜牧业生产中获得经济回报的私人利益。迫切需要采取旨在减少已观察到的权衡的力度和增加家庭经济收入的替代办法。
当前服务供应的限制
我们发现,在场地和景观尺度上,服务供给明显低于效率前沿形状所描述的最大可行水平。该地区的生物物理特征和目前的管理制度都促成了这种模式。土壤水分有效性的地形梯度造成的景观异质性导致了不同地点C种群(Jaramillo et al. 2003)、饲料(Trilleras et al. 2015)和物种丰富度(Balvanera et al. 2002)的重要差异。牧场维护不善可能导致饲料供应减少,特别是在生产力较高的地区,需要更频繁地燃烧以促进牧草生长和防止林木入侵(Trilleras et al. 2015)。此外,保护区内的原生林中C储量高于保护区外的原生林,可能是由于林产品的提取和放牧导致森林退化(Burgos and Maass 2004, Cohen Salgado 2014)。其他热带干旱林区也存在类似的退化问题(Chaturvedi et al. 2012, Ribeiro et al. 2015)。
服务对自然干扰或人为干预的响应时间滞后也可以解释供应水平的低效。当两种服务进行权衡时,达到首选服务的最大水平通常意味着与这些服务所依赖的生物物理基础的重新配置相关的时间延迟(Cavender-Bares et al. 2015)。牧草产量可能就是这样,随着牧场年龄的增长,牧草产量会增加(Trilleras等,2015年)。在人类干扰下通过森林更新恢复C种群的过程中,滞后时间要长得多:平均需要66年才能达到与保护森林相当的价值(Poorter et al. 2016)。
不同政府项目之间历来缺乏协调,导致土地管理目标相互冲突,这也可能阻碍实现可持续发展。养牛计划鼓励森林转变为牧场(Maass et al. 2005, Griscom和Ashton 2011)。另一方面,目前的环境项目,如环境服务付费(PES)或建立保护区,限制了资源的使用,仍然提倡“不接触”的管理方式。在该地区同时和不协调地执行这些政策鼓励农民发展低效的管理做法。
通过替代管理策略增加服务供应
模拟结果表明,在选择以增加服务供给为目标的管理策略时,应考虑景观转换的程度。在低砍伐率下,通过演替使次生林完全恢复使服务供应更接近效率边界。该地区较高的森林恢复率使这种替代方法成为可能(Mora等人,2015年,Poorter等人,2016年)。然而,次生林通常被再次清除以建立牧场。像REDD+这样的金融机制,明确说明了避免砍伐森林带来的C值节约和生物多样性效益(Phelps等人,2012年),可以通过提供替代家庭收入来鼓励次生林的再生(Borrego和Skutsch, 2014年)。
相比之下,在森林砍伐严重的情况下,迈向效率前沿的最佳选择似乎是提高牧场生产力。在Chamela-Cuixmala地区,目前通过更高频率的割伤和焚烧事件来提高牧草生产力(Trilleras et al. 2015),尽管这可能以对养分循环、土壤养分存量和饲料质量产生长期负面影响为代价(Lawrence et al. 2007, García-Oliva和Jaramillo 2011, Trilleras et al. 2015)。在热带地区增加牲畜产量的替代方法包括用本地草料树丰富牧草。这已被证明在具有类似气候条件的其他地区是成功的(Barrance等人,2003年,Dagang和Nair 2003年,Murgueitio等人,2011年),并正在进行测试(C. Gonzalez,个人沟通)或已经被Chamela-Cuixmala地区的一些农民实施(F. Mora,个人观察).此外,在森林砍伐严重的情况下,它的采用是有利的(Kaimowitz和Angelsen 2008)。此外,本地草料树的存在将增加C库,并在不影响饲料供应的情况下有利于牧场的树木多样性(Barrance et al. 2003, Dagang and Nair 2003),因此软化了它们的本地权衡。
当结合使用时,强化牧场生产和允许的次生林更新可以显著促进增加服务供应,减少森林维护和人民生计之间的权衡(Latawiec等,2015年)。有趣的是,我们的模拟表明,这并没有把服务供应带到效率边界。原始森林的管理制度也岌岌可危。当原始森林遭受木材提取和森林内的牛群放牧时,碳储量和物种丰富度都会减少(Chaturvedi et al. 2012, Ribeiro et al. 2015)。当牧场枯竭时,次生林和原生林是最重要的饲料来源,木材提取可能有助于家庭收入(Cohen Salgado 2014, Salmeróln 2015)。因此,防止或恢复森林退化是达到效率前沿的关键。REDD+正越来越多地解决森林退化的作用,农民认为其财政激励措施适合于预防和恢复森林退化(Skutsch等人,2015年)。
通过管理制度和技术的变化来处理这里所记录的权衡,面临着农民管理土地的方法随着时间的推移而根深蒂固的额外挑战。交流和促进新思想的采用是促进社会变革的种子(Rogers 2003)。目前正在通过讲习班、实地试验以及农民和科学家之间的合作项目,传播仍然不多的当地替代管理经验,以及分享关于可持续生态系统管理的科学研究结果。加强现有的社会网络和开放更多的信息交流空间似乎是满足农民需求和保障生计的途径,同时保持热带干燥森林在这样的条件下提供生态系统服务的能力,包括碳储存和生物多样性,不会被不可逆转地破坏。
结论
在效率前沿框架下,实现可持续性与将服务供给向效率前沿移动有关,该前沿的特定位置由利益相关者的需求、偏好和价值定义(Cavender-Bares等,2015年)。通过将这一框架应用到热带干燥森林景观的服务供应分析中,我们已经表明,尽管在用于牛的饲料生产和碳储量维持或物种保护之间存在强烈的权衡,但仍然有可能通过修改牧场和森林的管理,从目前的次优水平增加这些服务的供应。
允许完全恢复次生林,同时用原生草料树丰富牧场,是增加碳储量、促进景观规模生物多样性保护和维持农村生计的理想管理选择。为此目的,应优先给予经济奖励以维持目前的森林覆盖(从而维持C类种群和生物多样性保护),以及为其他管理做法提供技术支助。可以通过技术变革和财政激励的支持来解决热带地区为碳储存而进行的森林维护与生物多样性和农村生计之间普遍存在的权衡问题。在这里获得的见解有助于热带景观的可持续性。
致谢
这里发展的想法来自于过去15年与许多在Chamela-Cuixmala地区工作的同事进行的跨学科合作,这种综合是在国家生态分析与综合中心举办的讲习班所促进的。这项工作的资金支持来自PB的PAPIIT IN211114、SEP-CONACYT 2012-179045和SEP-CONACYT 2015-255544,以及AC的PAPIIT IN300813。我们感谢Chamela生物站(UNAM)对开展实地工作的支持,以及所有贡献了专业知识的当地利益相关方。
文献引用
阿查德,F., R.伯切尔,P.马约,H.-J。斯蒂比格,C.博德特,A.布林克,S.卡博尼,B. Desclée, F. Donnay, H. D. Eva, A. Lupi, R. raxi, R. Seliger,和D. Simonetti. 2014。确定1990 - 2010年热带森林砍伐率和相关碳损失。全球变化生物学20(8): 2540 - 2554。http://dx.doi.org/10.1111/gcb.12605
安德森-特谢拉,K. J. P. K. Snyder, T. E. Twine, S. V. Cuadra, M. H. Costa和E. H. DeLucia。2012.美洲自然和农业生态区的气候调节服务。自然气候变化beplay竞技2(3): 177 - 181。http://dx.doi.org/10.1038/nclimate1346
P.巴尔瓦涅拉,A.卡斯蒂略,M. J. Martínez-Harms。2011.季节性干燥热带森林的生态系统服务。259 - 277页在R.迪佐,H. S.杨,H. A.穆尼和G.塞巴洛斯,编辑。季节性干燥热带森林:生态与保护.美国华盛顿特区岛。http://dx.doi.org/10.5822/978-1-61091-021-7_15
P. Balvanera, E. Lott, G. Segura, C. Siebe和A. Islas. 2002。墨西哥热带干燥森林β-多样性的模式。植被科学杂志13(2): 145 - 158。http://dx.doi.org/10.1111/j.1654-1103.2002.tb02034.x
Barrance, A. J. Flores, E. Padilla, J. E. Gordon, K. Schreckenberg. 2003。洪都拉斯南部干旱地区的树木和农业I:农夫树管理实践。农林复合经营系统59:97 - 106。http://dx.doi.org/10.1023/A:1026347006022
比尔,1973年出生。树木密度对昆士兰西南mulga地区牧草和树木成分产量的影响(金合欢aneuraf . Muell)擦洗。热带草原7(1): 135 - 142。
Borrego, A.和M. Skutsch. 2014。估算导致热带干燥森林退化的活动的机会成本:对REDD+的影响。生态经济学101:1-9。http://dx.doi.org/10.1016/j.ecolecon.2014.02.005
布朗,C. J.和P. J.芒比。2014。渔业与生态系统功能保护之间的权衡是由管理策略确定的。生态学与环境前沿“,12(6): 324 - 329。http://dx.doi.org/10.1890/130296
布洛克,2002一个.科迪亚elaeagnoides直流。(紫草科)。Barcino。151 - 153页在F. A.诺格拉,J. H.维加,A. N. García-Aldrete和M.克萨达,编辑。历史自然德查米拉.国立大学研究院Biología,国立大学Autónoma de México, México D.F, México。
布洛克,2002b.Jacaratia墨西哥答:直流。(番木瓜科)。Bonete。155 - 157页在F. A.诺格拉,J. H.维加,A. N. Garcóa-Aldrete和M.克萨达,编辑。历史自然德查米拉.国立大学研究院Biología,国立大学Autónoma de México, México D.F, México。
Burgos, A.和J. M. Maass. 2004。墨西哥西部热带干燥森林地区与土地使用有关的植被变化。农业、生态系统与环境104(3): 475 - 481。http://dx.doi.org/10.1016/j.agee.2004.01.038
A.卡萨斯,S.兰格尔-兰达,I.托雷斯,E. Peréz-Negrón, L. Solís, F.帕拉,A.德尔加多,J.布兰卡斯,B. Farfán-Heredia,和A.莫雷诺-卡尔斯。2008.Tehuacán-Cuicatlán谷,México的植物资源就地管理和保护:一种民族植物学和生态学的方法。页1 - 25在U. De Albuquerque和M. Alves-Ramos,编辑。民族植物学的当前话题.研究路标,印度喀拉拉邦。
A.卡斯蒂略,A. Magaña, A.普贾达斯,L. Martínez, C. Godínez。2005.了解农村人口与生态系统的相互作用:墨西哥热带干燥森林的一个案例研究。生态系统八(6):630 - 643。http://dx.doi.org/10.1007/s10021-005-0127-1
Cavender-Bares, J., S. Polasky, E. King和P. Balvanera. 2015。评估生态系统服务权衡的可持续性框架。生态和社会20(1): 17。http://dx.doi.org/10.5751/es-06917-200117
查图维迪,R. K., A. S. Raghubanshi, J. S.辛格,2012。放牧和采伐对热带干旱森林幼树多样性、新生和碳积累的影响森林生态与经营284:152 - 162。http://dx.doi.org/10.1016/j.foreco.2012.07.053
科恩·萨尔加多,2014年。哈利斯科的查米拉-翠克斯马拉葡萄酒aledaño葡萄酒珍藏Biósfera.二元同步通信。论文。国立大学Autónoma de México,莫雷里亚,México。
科特勒,E. Durán和C.西贝。2002。Caracterización morfo-edafológica我们是热带地区的节日。17 - 79页在F. A.诺格拉,J. H.维加,A. N. García-Aldrete和M.克萨达,编辑。历史自然德查米拉.国立大学研究院Biología,国立大学Autónoma de México, México D.F, México。
戴刚,戴刚。2003。中美洲的森林对口研究和采用:最近的发现和对未来方向的建议。农林复合经营系统59:149 - 155。http://dx.doi.org/10.1023/A:1026394019808
de Albuquerque, up, L. de H. C. Andrade和A. C. O. de Silva, 2005。季节性干燥森林植物资源的利用(巴西东北部)。Acta神物铺子Brasilica19:27-38。http://dx.doi.org/10.1590/S0102-33062005000100004
杜兰,e . 2004。在哈利斯科的查米拉,这里有树木和风景.论文。国立大学Autónoma de México, México d.f., México。
Durán, E. P. Balvanera, E. Lott, G. Segura, A. Pérez-Jiménez, Á。Íslas, M. Franco, 2002。Estructura, composición y dinámica德拉vegetación。443 - 472页在F. A.诺格拉,J. H.维加,A. N. García-Aldrete, m .克萨达,编辑。历史自然德查米拉.国立大学研究院Biología,国立大学Autínoma de México, México D.F, México。
Durán, E. J. Meave, E. J. Lott, G. Segura. 2006。墨西哥热带落叶林景观层次的结构和树木多样性格局。Boletín la Sociedad Botánica de México79:43-60。
García-Oliva, F., A. Camou, J. M. Maass. 2002。El clima de la region de la costa del Pacífico mexicano。页3 - 10在F. A.诺格拉,J. H.维加,A. N. García-Aldrete和M.克萨达,编辑。历史自然德查米拉.墨西哥国立大学研究院Biología Autónoma de México
García-Oliva, F.和V. J. Jaramillo. 2011。季节性干旱热带森林的人为改造对生态系统生物地球化学过程的影响。159 - 172页在R.迪佐,H. S.杨,H. A.穆尼和G.塞巴洛斯,编辑。季节性干燥热带森林:生态与保护.岛/资源经济中心,华盛顿特区,美国。http://dx.doi.org/10.5822/978-1-61091-021-7_10
格里特森,J.罗萨莱斯,A.莫雷诺和D.卢埃特,2007。Ganadería我们的社会:我们的系统是生产力的,我们的系统是Sierra的Manantlán。页面27-52在L. M. Martínez里维拉和P. R. W.格里特森编辑。Estado real y perspective de la ganadería extensiva en la Sierra de Manantlán, en el occidente de México.瓜达拉哈拉大学,瓜达拉哈拉,México。
吉尔罗伊,J. J.伍德考克,F. A.爱德华兹,C.惠勒,C. A.梅迪纳·乌里韦,T.豪加森,D. P.爱德华兹。2014。优化热带农业景观的碳储存和生物多样性保护。全球变化生物学20(7): 2162 - 2172。http://dx.doi.org/10.1111/gcb.12482
高泰利,n.j.和R. K.科尔威尔,2001。生物多样性量化:物种丰富度测量和比较的程序和陷阱。生态学通讯4(4): 379 - 391。http://dx.doi.org/10.1046/j.1461-0248.2001.00230.x
H. P. Griscom和M. S. Ashton, 2011。中美洲干旱热带森林的恢复:模式和过程综述。森林生态与经营261(10): 1564 - 1579。http://dx.doi.org/10.1016/j.foreco.2010.08.027
哈维,C. A.和W. A.哈伯,1998。哥斯达黎加牧场的残树与生物多样性保护。农林复合经营系统44:37 - 68。http://dx.doi.org/10.1023/A:1006122211692
国际农业发展基金。2004.畜牧业服务与穷人:一项全球倡议。收集、协调和分享经验.国际农业发展基金,意大利罗马。(在线)网址:https://www.ifad.org/documents/10180/831c52c2-2ee2-4ff0-8d59-6aef6aa6dc97
V. J.贾拉米略,J. B.考夫曼,L. Rentería-Rodríguez, D. L.卡明斯,L. J.埃林森。2003。墨西哥热带干燥森林景观中的生物量、碳和氮库。生态系统6(7): 609 - 629。http://dx.doi.org/10.1007/s10021-002-0195-4
D. Kaimowitz和A. Angelsen. 2008。畜牧业集约化能帮助拯救拉丁美洲的热带森林吗?可持续林业杂志27(1 - 2): 6日到24日。http://dx.doi.org/10.1080/10549810802225168
King, E. G. J. Cavender-Bares, P. Balvanera, T. H. Mwampamba和S. Polasky. 2015。生态系统服务和利益相关者偏好的权衡:评估冲突、障碍和机会。生态和社会20(3): 25。http://dx.doi.org/10.5751/ES-07822-200325
兰宾,E. F., H. J.盖斯特,E.麻风病。2003。热带地区土地利用和土地覆盖变化动态。《环境与资源年报》28(1): 205 - 241。http://dx.doi.org/10.1146/annurev.energy.28.050302.105459
Latawiec, A. E., B. B. N. Strassburg, P. H. S. Brancalion, R. R. Rodrigues, T. Gardner. 2015。为热带农业景观的大规模修复创造空间。生态学与环境前沿“,13(4): 211 - 218。http://dx.doi.org/10.1890/140052
劳伦斯,D. D 'Odorico, L. Diekmann, M. DeLonge, R. Das和J. Eaton, 2007。森林砍伐后的生态反馈可能造成热带干燥森林灾难性的生态系统转变。美国国家科学院院刊104(52): 20696 - 20701。http://dx.doi.org/10.1073/pnas.0705005104
Lebrija-Trejos, E. F. Bongers, E. A. Pérez-García和J. A. Meave. 2008。农业转轨后干旱热带落叶林的演替变化与恢复力。《40(4): 422 - 431。http://dx.doi.org/10.1111/j.1744-7429.2008.00398.x
Lebrija-Trejos, E, E. A. Pérez-García, J. A. Meave, L. Poorter, F. Bongers. 2011。墨西哥热带干燥森林次生演替过程中的环境变化。热带生态学杂志27(5): 477 - 489。http://dx.doi.org/10.1017/s0266467411000253
莱彻,G。,j·r·斯基r . l . Chazdon n .诺顿美国j·赖特,j . a . Meave e·a·Perez-Garcia r·穆尼奥斯e . Romero-Perez安德拉德,j·l·安德拉德p . Balvanera j . m . Becknell t . v .便当,r . Bhaskar f . Bongers诉Boukili p h . s . Brancalion r·G·塞萨尔·d·a·克拉克·b·克拉克·d·克雷文,a . DeFrancesco j . m . Dupuy称:"现在b . Finegan e . Gonzalez-Jiminez j·s·霍尔,k . e .危害j·l·Hernandez-Stefanoni p . Hietz d . Kennard Killeen t·j·s·G·劳伦斯,e·e·Lebrija-TrejosM. Lohbeck, M. Martínez-Ramos, P. E. S. Massoca, R. C. G. Mesquita, F. Mora, R. Muscarella, H. Paz, F. Pineda-García, J. S. Powers, R. quesda - monge, R. R. Rodrigues, M. E. Sandor, L. Sanaphre-Villanueva, E. Schüller, N. G. Swenson, A. Tauro, M. Uriarte, M. van Breugel, O. Vargas-Ramírez, R. A. G. Viani, A. L. Wendt, G. Bruce Williamson. 2015。环境梯度与生境专业化演化:14个新热带森林样地的试验案例。《生态学杂志》103:1276 - 1290。http://dx.doi.org/10.1111/1365-2745.12435
马斯,J. M. P.巴尔瓦内拉,A.卡斯蒂略,G. C.戴利,H. A.穆尼,P.埃利希,M.克萨达,A.米兰达,V. J.贾拉米略,F. García-Oliva, A. Martínez-Yrizar, H.科特勒,J. López-Blanco, A. Pérez-Jiménez, A. Búrquez, C.蒂诺科,G.塞巴洛斯,L.巴拉扎,R.阿亚拉,J.萨鲁坎。2005。热带干燥森林的生态系统服务:来自墨西哥太平洋海岸长期生态和社会研究的见解。生态和社会10(1): 17。(在线)网址://www.dpl-cld.com/vol10/iss1/art17/http://dx.doi.org/10.5751/es-01219-100117
马斯、C. F.乔丹和J. Sarukhan, 1988。不同管理技术下热带季节性农业生态系统的土壤侵蚀和养分流失。应用生态学杂志25(2): 595 - 607。http://dx.doi.org/10.2307/2403847
2010。Martínez-HarmsCuantificación我们的服务之路ecosistémicos墨西哥之路:暗含的信息conservación y manejo.论文。国立大学Autónoma de México,莫雷里亚,México。
Martinez-Ramos, M, l . Barraza p . Balvanera j·贝尼特斯,f . Bongers a·卡斯蒂略a·卡隆·g·伊瓦拉,h . Paz-Hernandez a . Perez-Jimenez M, d . r . Perez-Salicrup g . a . sanchez - azofeifa告诉本网络记者,j . e . Schondube k碎石机,j·a·迪亚兹k . Boege e·德尔瓦尔·M·e·f·Carrillo Suazo-ortuno, l·d·阿维拉——M·阿尔瓦雷斯M·卡诺j·卡斯蒂略,o·查维斯,e·德拉佩纳a .科尔索c . Godinez a·p·戈麦斯a·冈萨雷斯,b . Fuentealba w . Gudino o·埃尔南德斯,M . Lobeck c . ManriqueS.马扎-比利亚洛沃斯,M. Méndez, F.莫拉-阿迪拉,C.穆恩奇,P.克里斯蒂娜,L. Pinzón, M. E. Páramo, F.皮内达,A. Ricaño, M.罗查,J. Rodríguez-Velázquez, N.施罗德,J.特里莱拉-莫塔,M.范·布勒格尔,P.范·德斯利恩,E.维拉,和I. Zermeño。2012.Manejo de bosques tropical: científicas para la conservación, restauración y aprovechamiento de生态系统emas en paisajes乡村。Investigación环境,科学Política Pública4(2): 111 - 129。
Martínez-Yrízar, A. J.萨鲁坎,A. Pérez-Jimínez, E. Rincón, J. M.马斯,A. Solís-Magallanes, L.塞万提斯,1992。哈利斯科海岸的热带落叶林的地上植物体,México。热带生态学杂志8(01): 87 - 96。http://dx.doi.org/10.1017/s0266467400006131
Maza-Villalobos, S., P. Balvanera和M. Martínez-Ramos。2011.热带干旱森林从废弃牧场的早期再生:时间序列和动态方法的对比。《43(6): 666 - 675。http://dx.doi.org/10.1111/j.1744-7429.2011.00755.x
麦基安,2000年。公共属性:它是什么,它有什么好处,是什么使它起作用?页面27-55在c·c·吉布森,m·a·麦基恩,e·奥斯特罗姆,编辑。人与森林:社区、机构和治理.麻省理工学院出版社,剑桥,马萨诸塞州,美国。
F. Mora, M. Martínez-Ramos, G. Ibarra-Manríquez, A. Pérez-Jimínez, J. Trilleras, P. Balvanera. 2015。通过动态方法测试时间序列:时间和地点对热带干燥森林演替的影响。《38 - 48。47 (1):http://dx.doi.org/10.1111/btp.12187
Murgueitio, E, Z. Calle, F. Uribe, A. Calle和B. Solorio. 2011。原生树木和灌木用于热带牛牧场土地的生产性恢复。森林生态与经营261(10): 1654 - 1663。http://dx.doi.org/10.1016/j.foreco.2010.09.027
奥克萨宁,J., F. G.布兰切特,R.金特,P.勒让德,P. R.明钦,R. B.奥哈拉,G. L.辛普森,P.索利莫斯,H. H.史蒂文斯,H.瓦格纳。2011。素食:社区生态套餐.R包版本2.0-2。R基金会,维也纳,奥地利。(在线)网址:https://cran.r-project.org/
菲尔普斯,J. E. L.韦伯和W. M.亚当斯,2012。生物多样性是减少森林碳排放政策的共同利益。自然气候变化beplay竞技2:497 - 503。http://dx.doi.org/10.1038/nclimate1462
Pineda-García, F., H. Paz,和F. C. Meinzer. 2013。热带干旱森林早期和晚期次生演替树种的抗旱性:木质部抗栓塞性、边材贮水和叶片脱落之间的相互作用。《植物、细胞与环境36(2): 405 - 418。http://dx.doi.org/10.1111/j.1365-3040.2012.02582.x
Polasky, S, E. Nelson, J. Camm, B. Csuti, P. Fackler, E. Lonsdorf, C. Montgomery, D. White, J. Arthur, B. Garber-Yonts, R. Haight, J. Kagan, A. Starfield, C. Tobalske. 2008。把东西放在哪里?空间土地管理以维持生物多样性和经济回报。生物保护141(6): 1505 - 1524。http://dx.doi.org/10.1016/j.biocon.2008.03.022
要隘,L f . Bongers t . m .助手,a . m . Almeyda摩尔诺,p . Balvanera j . m . Becknell诉Boukili p h . s . Brancalion e . n . Broadbent r . L . Chazdon d·克雷文j . s . de Almeida-Cortez g·a·L·卡布拉尔b·h·j·德容j . s . Denslow d·h·削弱s . j . DeWalt j . m . Dupuy称:"现在s·m·杜兰·m·m·圣埃斯皮里图m . c . Fandino r·g·塞萨尔j·s·霍尔,j·L·Hernandez-Stefanoni c . c . Jakovac a b,琼奇拉d . Kennard s . g .莱彻J.-C。Licona, m . Lohbeck e . Marin-Spiotta m . Martinez-Ramos p . Massoca j . a . Meave r . Mesquita f•莫拉r·穆尼奥斯r . Muscarella y . r . f . Nunes s Ochoa-Gaona a . a . de Oliveira e . Orihuela-Belmonte m . Pena-Claros e·a·Perez-Garcia d . Piotto j . s .权力,j . Rodriguez-Velazquez即Romero-Perez, j . Ruiz j·g . Saldarriaga sanchez - azofeifa告诉本网络记者,n . b . Schwartz m·k。施n g·斯文森,m·托莱多m . Uriarte m . van Breugel h . van der细胞膜,m·d·m·维罗索h·f·m·背心A. Vicentini, i.c.g.维埃拉,T. V. Bentos, G. B. Williamson, D. M. A. Rozendaal. 2016。新热带次生林生物量恢复力。自然530(7589): 211 - 214。http://dx.doi.org/10.1038/nature16512
Portillo-Quintero, C., A. Sanchez-Azofeifa, J. Calvo-Alvarado, M. Quesada, m.m. do Espirito-Santo。2015.热带干燥森林在新热带地区生物多样性、碳和水保存方面的作用:吸取的教训和可持续管理的机会。区域环境变化15(6): 1039 - 1049。http://dx.doi.org/10.1007/s10113-014-0689-6
Quijas, s . 2012。Diversidad vegetal y generación de servicios ecosistémicos一个不同的escalas espaciales.论文。国立大学Autónoma de México,莫雷里亚,México。
R开发核心团队,2015。R:用于统计计算的语言和环境.R统计计算基金会,维也纳,奥地利。(在线)网址:http://www.R-project.org/
Raudsepp-Hearne, C. G. D. Peterson和E. M. Bennett, 2010。生态系统服务包用于分析不同景观的权衡。美国国家科学院院刊107(11): 5242 - 5247。http://dx.doi.org/10.1073/pnas.0907284107
Rendón-Carmona, H., A. Martínez-Yríar, P. Balvanera和D. Pérez-Salicrup。2009.墨西哥热带干燥森林木本物种的选择性砍伐:使用和保护之间的不相容。森林生态与经营257(2): 567 - 579。http://dx.doi.org/10.1016/j.foreco.2008.09.031
Ribeiro, E. M. S, V. Arroyo-Rodríguez, B. A. Santos, M. Tabarelli和I. R. Leal. 2015。长期的人为干扰导致巴西Caatinga植被的生物贫瘠。应用生态学杂志52(3): 611 - 620。http://dx.doi.org/10.1111/1365-2664.12420
Rodríguez, J. P. T. D.比尔德,E. M.班尼特,G. S.卡明,S. J.科克,J.阿加德,A. P.多布森,G. D.彼得森。2006。跨越空间、时间和生态系统服务的权衡。生态和社会11(1): 28。(在线)网址://www.dpl-cld.com/vol11/iss1/art28/http://dx.doi.org/10.5751/es-01667-110128
罗杰斯,e。m。2003。创新的扩散.第五版。免费,纽约,纽约,美国。
瑞吉斯,A.沃斯克,M.科尔特莱宁,R.阿尔克玛德,C. J. E. Schulp. 2013。东欧生态系统服务的权衡分析。生态系统服务4:82 - 94。http://dx.doi.org/10.1016/j.ecoser.2013.04.002
Salmerón, o.u. 2015。英勇económico我们是哈利斯科夏米拉热带雨林的森林之神.论文。国立大学Autónoma de México,莫雷里亚,México。
Sánchez-Azofeifa, G. A.和C. portilo - quintero。2011.新热带季节干燥热带森林变化的范围和驱动因素。页面45-57在R.迪佐,H. S.杨,H. A.穆尼和G.塞巴洛斯,编辑。季节性干燥的热带森林。生态学和保护.美国华盛顿特区岛。http://dx.doi.org/10.5822/978-1-61091-021-7_3
Sánchez-Azofeifa, G. A., M. Quesada, P. Cuevas-Reyes, A. Castillo, G. Sánchez-Montoya。2009.墨西哥Chamela-Cuixmala生物圈保护区影响区域的土地覆盖和保护。森林生态与经营258(6): 907 - 912。http://dx.doi.org/10.1016/j.foreco.2008.10.030
影子,2002年出版。Tierra, trabajo y ganado en la región北哈利斯科:una historia agraria de Villa Guerrero,哈利斯科,1600-1980.El Colegio de Michoacán AC,萨莫拉,México。
瑟塞利,J.和S.纳伊姆,2012。小农休耕区林下乔灌木与牧草的关系反映了生态系统服务的相互作用。农林复合经营系统87(2): 451 - 464。http://dx.doi.org/10.1007/s10457-012-9566-9
M. Skutsch, A. Borrego, L. Morales-Barquero, J. Paneque-Gálvez, M. Salinas-Melgoza, m.i. Ramírez, D. Pérez-Salicrup, D. Benet, S. Monroy,和Y. Gao. 2015。农村社区在促进REDD +下森林景观转变的潜力方面的机会、制约因素和看法:来自墨西哥的案例研究。国际林业审查17 (SI): 65 - 84。http://dx.doi.org/10.1505/146554815814669025
索托,j.c. 2009。植物útiles de la cuenca del Balsas。277 - 310页在G.塞巴洛斯,L. Martínez, A. García, E.埃斯皮诺萨,J.贝扎里和R.迪佐,编辑。多样化,我们的任务áreas优先任务conservaciín de Selvas Secas del Pacíico de México.文化Fondo de culture Económica, México, D.F.
H.塔利斯,P. Kareiva, M. Marvier和A. Chang. 2008。一个支持实际保护和经济发展的生态系统服务框架。美国国家科学院院刊105(28): 9457 - 9464。http://dx.doi.org/10.1073/pnas.0705797105
特里莱拉斯,J. M., V. J.贾拉米略,E. V.维加,P. Balvanera. 2015。墨西哥西部热带干旱地区畜牧业管理对牧场生态系统服务供给的影响。农业、生态系统与环境211:133 - 144。http://dx.doi.org/10.1016/j.agee.2015.06.011
