

版权所有©2012作者。由韧性联盟授权在此发布。
去pdf本文的版本
去pdf本文的版本
以下是引用这篇文章的既定格式:
埃希霍恩,M., K. Johst, R. Seppelt, M. Drechsler. 2012。基于模型的掠食性鸟类与风力涡轮机碰撞风险估计。生态和社会 17(2): 1。
http://dx.doi.org/10.5751/ES-04594-170201
研究
埃希霍恩,M., K. Johst, R. Seppelt, M. Drechsler. 2012。基于模型的掠食性鸟类与风力涡轮机碰撞风险估计。生态和社会 17(2): 1。
http://dx.doi.org/10.5751/ES-04594-170201
基于模型的捕食性鸟类与风力涡轮机碰撞风险估计
1海姆霍兹环境研究中心,2UFZ-Helmholtz环境研究中心
摘要
风能等可再生能源的发展是减缓气候变化的一种有前途的方式。beplay竞技由于风力涡轮机有与旋翼叶片碰撞的风险,对当地鸟类的数量有负面影响,特别是对迅猛龙,如红鸢(Milvus Milvus).对这些影响缺乏适当的评估工具。为了弥补这一差距,我们开发了一个基于主体的空间显式模型,模拟红鸢在不同土地使用类型的景观中围绕其巢穴的觅食行为。我们确定了红风筝与涡轮机的碰撞风险作为一个函数的风力涡轮机和aerie之间的距离和其他参数。影响函数包括种属觅食行为和景观结构的协同效应。碰撞风险随距离的增加呈指数下降。这种下降的强度取决于迅猛龙的觅食行为,它避开风力涡轮机的能力,以及该地区的平均风速。通过仿真模型估计的碰撞风险在现场观测值的范围内。推导的碰撞函数表明,碰撞风险可以描述为风力发电机与猛禽巢穴之间距离的聚集函数。这使得(现有或计划中的)风力涡轮机的生态影响可以根据其空间位置进行简单和快速的评估。 Furthermore, it implies that minimum buffer zones for different landscapes can be determined in a defensible way. This modeling approach can be extended to other bird species with central-place foraging behavior. It provides a helpful tool for landscape planning aimed at minimizing the impacts of wind power on biodiversity.
关键词:碰撞风险;觅食行为;影响评估;基于单独的模型;Milvus Milvus;红色的风筝;空间模型;风力发电
介绍
欧洲鼓励扩大可再生能源生产,以缓解气候变化。beplay竞技在许多欧洲国家,这需要增加新的陆上风力发电厂的数量。土地稀缺和土地使用冲突使这成为一项具有挑战性的任务(Ohl和Eichhorn 2010年)。确定风力涡轮机(WTs)的合适地点不仅需要满足风电场运营商的需求,还需要考虑风力发电的相关不利影响,例如对人类健康和野生动物的影响。生物学家强调了对鸟类种群的潜在影响,如干扰和迁移(Percival 2000, 2005, Drewitt and Langston 2006, Larsen and Guillemette 2007, Pearce-Higgins et al. 2009)。碰撞导致高死亡率,特别是在WTs附近观察到的迅猛龙(Erickson等,2001年,Hunt 2002年,Barrios和Rodriguez 2004年,Smallwood和Thelander 2007年,Lekuona和Ursúa 2007年,de Lucas等,2008年,Krijgsveld等,2009年)。
目前的研究集中在红风筝的风险(Milvus Milvus)与wt相撞。这种掠食性鸟类属于被旋翼叶片撞击频率最高的物种(Mammen和Dürr 2006,勃兰登堡国家鸟类保护中心(BSBCC) 2010)。这些异常高的碰撞率的原因还没有被完全理解,但一些事实是重要的。风力涡轮机通常位于开放的农业景观,这是红风筝的主要栖息地。此外,迅猛龙在这些景观中的测距(飞行)行为助长了碰撞死亡。此外,红鸢会被吸引到更靠近涡轮机的区域,那里通常有较高的猎物丰度(Dürr 2009, jedike 2010)。
为了减少小波对红鸢等迅猛龙的影响,必须仔细选择小波的地点。在拥有世界上大约一半红风筝人口的德国,20世纪90年代出现了风力发电场不受控制的增长。从那时起,联邦各州倾向于通过区域规划来指导WTs的分配。结果,wt被集中在某些领域(将来也会如此),而被排除在其他领域之外。然而,正如Madders和Whitfield(2006)所指出的,规划决策往往完全基于主观判断,这是因为决定碰撞风险的过程信息的缺乏。
为这项评估建立了不同的科学方法。一种可能的方法是空间建模(描述活动范围和行为),通过预测感兴趣物种的活动,在景观尺度上确定鸟类最敏感的区域(Williams et al. 1996, McGrady et al. 1997, McLeod et al. 2003, Madders and Whitfield 2006)。Madders和Whitfield(2006)将这种空间模型列为影响评估的良好起点,因为它们具有成本效益,并且可以识别关键位置。然而,由于没有考虑物种的觅食行为和飞行模式,因此很难得出关于由此产生的碰撞风险的结论。第二种方法是使用碰撞风险模型(CRM),该模型首先由Tucker(1996)开发,随后由Biosis Research(2003)和Podolsky(2003, 2005)进一步开发。这些研究明确地考虑了WTs和鸟类的相互作用。该模型基于数学方程,根据在拟议风电场区域观察到的鸟类数量和飞行时间在旋翼扫掠体积中的比例,将经验数据纳入其中。此外,考虑了WT参数和目标鸟的大小、飞行方向和飞行速度(Madders和Whitfield, 2006)。所谓的“频带碰撞风险模型”(Band et al. 2007)是在英国广泛使用的CRM。
空间模型能够预测一个物种是否居住在某一区域,但不能提供关于碰撞风险的信息。相比之下,crm能够预测碰撞风险,但是为特定的风电场项目设计的,需要大量基于实地调查的输入数据(Band et al. 2007)。
在本研究中,考虑到红风筝的飞行和觅食行为以及景观的空间结构,将这两种方法的各个方面纳入到一个空间显式仿真模型中。我们研究了碰撞风险是如何取决于WT和红鸢巢穴之间的距离。Eichhorn和Drechsler(2010)在景观规划背景下提出了这样的影响函数(IF)。它以空间显式但聚集的方式描述了碰撞风险,因此是评估候选WT站点碰撞风险的一种快速而实用的工具。然而,Eichhorn和Drechsler(2010)使用的IF仅以定性的方式预测影响,并基于一些临时假设。因此,其实际形态及其对物种和景观特征的依赖性尚不清楚。
为了深入了解特定景观中物种行为的大规模效应,基于主体或个体的模型(ABM/IBM)非常适合(houston, et al. 1988, DeAngelis and Mooij 2005, Grimm and Railsback 2005)。我们开发了一个ABM,作为aery - wt距离的函数,用于碰撞风险的定量预测,并将其与Band CRM的具体发现相结合。在我们的ABM中,“代理”代表单个迅猛龙个体,其行为取决于栖息地需求和景观特征。为了确定可变aery - wt距离下的碰撞概率,利用ABM进行了虚拟实验。为了验证ABM,采用了面向模式的方法(Wiegand等人,2003年,Grimm等人,2005年),在模拟输出和现场观测之间进行模型拟合练习。
方法
为了模拟红鸢的觅食飞行对景观结构的响应,该模型基于对红鸢行为的现有知识。这种基于主体的方法通常允许评估特定物种对特定时间和/或空间景观属性的脆弱性(例如,Verboom等人1991年,Johst等人2001年,Wichmann等人2004年,Rodríguez等人2006年,Bauer等人2008年)。对于像红鸢这样的中心觅食者来说,关注的属性是小WT在巢穴附近的位置。该脆弱性由猛禽与该小波的碰撞风险来表示。我们应用该建模方法来预测小波对西萨克森州地区红鸢的影响。
模型描述
模型描述遵循ODD(概述、设计概念、细节)协议,用于描述个体和基于代理的模型(ABM/IBM) (Grimm et al. 2006)。目的
ABM的目的是确定中心觅食鸟类(这里是红鸢)的年死亡率,作为其巢穴和WT之间距离的函数。为此,ABM考虑了该物种的觅食行为和景观特征。该模型包含了理解鸟类和WTs在觅食过程中相互作用的必要过程。
基于建模结果,我们推导出一个将碰撞风险量化为aerie-WT距离函数的冲击函数,该函数与Eichhorn和Drechsler(2010)的结果相当。然后,我们探索这种聚合方法在多大程度上可以捕获物种特定觅食行为和景观结构对碰撞风险的协同效应。
状态变量和尺度
ABM包括三种类型的实体:构成景观的网格单元、Red Kite和WT(见表1)。在本研究中,我们考虑了一个与德国西萨克森州(West Saxony)土地利用模式类似的模型景观。我们选择这个地区是因为它位于红风筝全球分布的核心区域。
红风筝代理被设计成在觅食飞行中表现得像一只典型的迅猛龙。关于它的行为和对景观的反应的信息来自于我们自己的现场观察、专家访谈(Hötker,个人交流2008)和文献(Whitfield和Madders 2006b, Nachtigall 2008)。只有成对的雄性迅猛龙被考虑,因为它们在繁殖季节有更多的飞行活动。静息行为不被考虑,因为它不会增加碰撞风险。
WT是由它相对于位于模型世界中心的红鸢巢的空间位置和细胞的使用类型决定的。
大多数碰撞发生在繁殖季节,这时经常有觅食飞行来喂养雏鸟(Mammen和Dürr 2006, Dürr 2009)。红鸢的繁殖期为85天(4月至6月),这是一个模型运行的长度。因此,我们假设在这段时间内发生的碰撞充分描述了每年的碰撞风险。24小时的白天不仅包括觅食飞行,还包括休息时间和夜间活动。根据Nachtigall(2008)观察到的离巢时间,我们计算出了平均10.6小时/天的“活动期”(鸟活跃并因此离开巢的时间)。Nachtigall(2008)估计休息时间大约是“活动期”的50%,所以鸟每天飞行5.3小时,或450小时/85天。因此,一个模式年相当于飞行450小时。为了计算每次模型运行的时间,我们记录了飞行距离,并除以迅猛龙的飞行速度(表1)。
流程概述和调度
ABM基于三个运动过程(随机飞行、定向飞行和向前飞行)和两个事件过程(碰撞事件和捕获猎物)。在每个建模步骤的开始,鸟类对被占领的细胞和邻近细胞的栖息地条件进行采样。有三种情况是可能的:
- 被占领的细胞和所有邻近的细胞是相同的,并不是最差的栖息地质量,导致随机飞行过程。在这里,agent向前移动一个细胞,然后以一定的概率(1-pf)/2分别向右或向左飞一圈;
- 相邻细胞的栖息地质量不同,导致定向飞行过程。在这里,代理移动到生境质量最好的细胞之一,并以(1-)的概率进行决策pf)/2分别向右或向左飞一圈;而且
- 被占领的细胞是最差的栖息地质量,启动飞行向前程序,代理移动一个细胞(100米)向前。
如果猛禽穿过WT转子叶片所跨越的区域,而不主动试图避免打击,它将被叶片击中。通过确定各自的概率,将这些情况纳入ABM:
- 发生碰撞的基本条件是迅猛龙到达WT所在的区域(细胞)。
- 红风筝在WT旋翼所覆盖的高度(p跳频).取具有轮毂高度的WThh= 78米和转子长度r= 41 m,使用Mammen(2010)的经验数据,我们估计的概率p跳频= 0.34。
- 概率(prsa)猛禽经过受旋翼影响的区域。为了得出一个简单的估计,我们假设迅猛龙水平地飞到陆地表面,并与旋翼扫过的平面正交。假设猛禽飞行的高度区间是由WT的中心高度决定的(hh)加/减一个动叶长度(r).这就产生了水平方向上由单元宽度(w = 100 m)和垂直方向上由转子直径(2r)跨越的飞行窗口,从而产生了一个矩形面积为2rw的区域。转子叶片所跨的圆面积等于πr2.假设飞行窗口内飞行高度分布是均匀的,则猛禽飞过旋翼掠区的概率为:
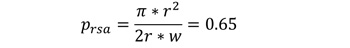
(1) - 鸟飞过旋翼时被击中的概率(即物理碰撞概率)p乐队)使用带碰撞风险模型的第二阶段计算(Band et al. 2007)。“这种可能性取决于鸟的大小(包括长度和翼展),涡轮叶片的宽度和间距,涡轮的转速,当然还有鸟的飞行速度”(苏格兰国家遗产(SNH) 2000年)。由苏格兰政府资助的自然保护机构SNH提供了一个计算模板(SNH 2010)。采用不同的桨叶螺距角值考虑了风速的不确定性,得到的值为p乐队在0.144到0.205之间。
- 迅猛龙识别威胁并主动避免碰撞的概率(pav).在大多数情况下,迅猛龙会扫描周围环境,并积极避免与基础设施发生碰撞。这种行为可以用回避概率来概括pav.估算规避概率是一项具有挑战性的任务(Chamberlain et al. 2006, Whitfield and Madders 2006a, Whitfield 2009)。Smales和Muir(2005)在澳大利亚的碰撞风险建模中使用了95%、98%和99%三个任意规避概率。Whitfield和Madders (2006b)利用西班牙北部13个风力发电场的几种掠食性鸟类的数据,经验估计规避概率在98%到100%之间,红鸢的可能值为99%。在目前的研究中,我们考虑了四个可能的水平pav: 98%、98.5%、99%、99.5%。
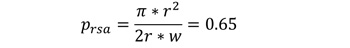
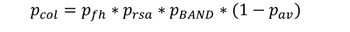 |
(2) |
捕获猎物的过程决定了红风筝在开始下一次觅食飞行之前捕获猎物并返回巢穴的时间点。这一过程使用Nachtigall(2008)提供的空降时间的频率分布参数化,考虑到约50%的空降时间用于飞行(见第一部分状态变量和尺度)。这种飞行时间的频率分布是离散的,它提供了在特定间隔内观察到飞行时间的概率。我们假设在每个时间区间内飞行时间是均匀分布的,从而得到飞行时间的连续概率密度函数(图1)。图2给出了经过上述步骤的ABM流程图。
相关的设计概念
特性转化
虚拟景观是随机生成的,假设不同生境类型的份额固定。此外,迅猛龙在飞行过程中的行为决定在很大程度上是随机的,而且取决于概率pf的概率密度函数和飞行持续时间的概率密度函数(图1)。模型运行10000次,对相关变量进行统计分析。
初始化
通过为每次模拟运行创建一个随机的视图来初始化模型视图。具有特定生境品质的细胞数目(总部= 0,1,2,3)是固定的,基于西萨克森地区的景观结构。Red Kite代理在位于模型世界中心的巢穴中初始化。WT实体初始化在一个随机选择的位置,受两个约束条件的限制。首先,它只能生长在适合的生境单元上,例如森林(总部= 0)被排除。其次,它位于与巢穴指定的距离上,距离从100米到500米不等,步距为200米,步距为500米,从500米到5000米不等。
输入
输入数据用于两个过程。首先,我们在参考地区西萨克森州观察到的土地使用类型的比例被用来生成虚拟景观。该地区的特点是开放农业用地比例高,森林面积比例小。从真实土地覆盖数据(ATKIS 2007)中提取的不同土地利用类型被归纳为具有不同生境质量水平的四类。草地和牧场的生境质量最高(总部= 3),占西萨克森州约11%的生境质量总部= 2(例如,农田)约占55%,生境质量总部= 1(例如居民点)约占4%,而森林面积的质量最低(总部= 0),约占景观面积的17%。
交互
红风筝代理与周围景观相互作用,景观的质量影响代理的飞行行为。第二个交互发生在红风筝和WT之间。如果满足碰撞事件过程中定义的条件(参见过程概述和调度部分),红风筝就会与WT发生碰撞。
模拟觅食行为的验证
我们通过比较模型的输出和经验行为数据来验证模型中的觅食行为。尽管红鸢的详细现场数据很少,但Nachtigall(2008)确定了与飞巢在一定距离级内的飞巢外驻留频率及其与飞巢的最大距离,并已用于验证。策略
有两个模型参数,主要控制红风筝代理的飞行行为:概率pf和最长停留时间T在飞行途中的手机上。由于其不确定性,似然值是通过面向模式的建模来估计的(Grimm et al. 2005)。的概率pf取值范围为0 ~ 1和停留时间T从1.2分钟到12分钟(Nachtigall,个人沟通,2009)。不同pf在0.1和T步骤1.2分钟,并系统组合pf 而且T因为所有值导致110个参数组合,因此称为“策略”。
最佳策略的确定
为了确定哪一种策略导致模型和经验行为之间的最佳吻合,我们为每种策略运行了10,000次模型,并记录了驻留频率x我迅猛龙的四个不同距离等级(我= 1,…,4)距巢穴:0 - 1公里(我= 1),介乎1至2公里(我= 2),介乎2至3公里(我= 3),及大于3km (我= 4)。Nachtigall(2008)确定了以下这些距离类别的频率:y1= 0.6,y2= 0.2,y3.= 0.15,y4雄性红鸢= 0.05。除了距离类,我们比较了模型的最大距离aerie(表示为k),值为l马克斯= Nachtigall(2008)在野外观测到的4500米。
相对偏差σ我(x)我从经验数据y我对于每个距离类(i),用
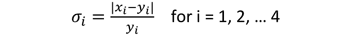 |
(3) |
各距离类的平均相对偏差σdistclas通过
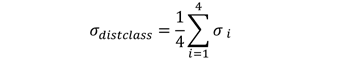 |
(4) |
相对偏差σmaxdist关于最大距离的计算方法是
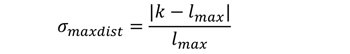 |
(5) |
通过最小化σdistclass和σmaxdist,我们确定了最符合实证数据的策略。
碰撞风险分析
对于提供与经验数据最吻合的模型的策略,我们确定了一组场景的碰撞风险。每种情况由aery - wt距离和规避概率的组合定义pav.aere - wt距离范围为100 ~ 5000 m,共11步,规避概率为100 ~ 5000 mpav从0.98到0.995的三个步骤(参见小节过程概述和调度),总共导致48个场景。为了找出最佳策略和48种情况,我们将该模型运行了1万次。对于每一个模型运行,生成一个与研究区域栖息地类型比例相同的随机景观,WT被随机放置在距离飞禽的指定距离,我们记录是否发生碰撞。然后,通过将碰撞事件的次数除以模型运行的次数,计算每个场景的碰撞风险。
结果
觅食行为
该模型中飞行行为的特征是向前飞行的概率和停留在细胞上的时间。对于110种不同的策略,每一种都由向前飞行的概率的组合定义pf和最长停留时间T,模型数据与经验数据之间的相对偏差(Nachtigall 2008)σ distclass而且σ maxdist计算方法详见上述“觅食行为的验证”小节。图3显示了模型拟合迅猛龙在不同距离上被观察到的频率的函数结果pf而且T.它的差异很大pf但只弱依赖于T.最小相对偏差σdistclass为0.17pf= 0.4,T= 12分钟。
模型拟合也应用到最大飞行距离的aerie (σ maxdist的函数pf而且T.类似于σ distclass在图3中,σ maxdist强烈的变化pf但只弱依赖于T.最小相对偏差σ maxdist的值pf= 0.3,T= 3.6分钟。
由模型推导出的两种策略符合(pf= 0.4,T= 12分钟和pf= 0.3,T= 3.6分钟)代表相似的水平pf(0.4 vs. 0.3),在T(12分钟vs. 3.6分钟)然而,如上所述,的水平T只能微弱地决定模型输出和观测值之间的偏差(Nachtigall 2008)。因此,两种策略都提供了令人满意的模型拟合,相对偏差小于20%。作为战略pf= 0.4,T= 12分钟表现稍好σ distclass而战略(pf= 0.3,T= 3.6分钟)表现更好σ maxdist,我们在分析碰撞风险时,两者都要考虑。
碰撞的风险
我们拟合的飞行行为模型揭示了两种与观测到的现场数据相对应的策略。对于这些策略,(pf= 0.4,T= 12分钟,图4b,d,和pf= 0.3,T= 3.6 min,图4a,c),进行碰撞风险分析,推导碰撞函数。图4显示了四种规避概率下,以年死亡率作为aerie-WT距离函数表示的碰撞风险pav以及两种不同的物理碰撞概率p乐队.正如预期的那样,碰撞风险随着aery - wt距离的增加而降低。曲线呈指数形状,因此碰撞风险在靠近空巢处迅速下降,在距离较远处下降较慢。这些影响函数表明指数依赖性(即exp(-距离))比钟形影响函数(即exp(-距离))更合适2))在我们之前的研究(Eichhorn和Drechsler 2010)中假设。
冲击函数一般取决于停留时间T,规避概率pav,物理碰撞概率p乐队.停留时间较短的飞行行为导致WT碰撞导致的年死亡率较高。规避概率的降低会增加碰撞风险。例如,在较短的空气- wt距离(<1500米)下,死亡率的差异可达0.4pav在0.98到0.995之间。的增加p乐队随着风速的增加,猛禽被旋翼叶片击中的概率会增加,这是因为WT旋翼叶片在适应更高风速时发生了几何变化(图4a,b vs.图4c,d)。
讨论
提高风能扩展可持续性的一个重要步骤是评估WTs对鸟类的碰撞风险对生态的影响。在这一评估中很少调查的一个方面是碰撞风险与鸟类巢穴和景观中WT之间的距离之间的关系。因此,我们开发了一个基于agent的模型,用来研究一种觅食食肉鸟类的这种关系。红鸢是濒危物种之一,被怀疑受到西隧的影响。基于agent的模型模拟了红鸢的觅食飞行模式,并预测了由于在离巢不同距离处与WT碰撞而导致的年死亡率。从这些模拟中,我们能够推导出一个所谓的碰撞函数,描述了年碰撞风险和aerie-WT距离之间的函数关系。它聚集了物种特有的觅食行为和景观结构(包括WT的存在)之间的相互作用。我们发现,碰撞风险随着鸟类巢穴与WT之间距离的增加呈指数下降。这种下降的强度敏感地依赖于实施的规避概率pav它指定了鸟主动避免被旋翼叶片击中的可能性和鸟的特定飞行行为(由参数给出)T而且pf,见表1)。
因此,我们计算了覆盖Whitfield和Madders (2006b)给出的值的四种不同回避概率的冲击函数,并演示了碰撞风险的不确定性范围。红鸢的模拟飞行行为是通过比较模型输出与现场数据的频率观察鸟在特定的距离等级和它的最大飞行距离的巢穴验证的。模拟频率与观测频率的相对偏差小于20%。需要注意的是,在目前的模型中,红风筝的飞行行为取决于实际的环境条件,包括一个随机的组件,而不是内存。事实上,猛禽更喜欢巢穴周围的某些区域,因为猛禽被期望能够记住好的狩猎区域(Nachtigall,个人交流,2009)。然而,缺乏足够的实证数据。更好地了解鸟类觅食行为的一种可能的实验方法是,例如GPS卫星遥测,它已成功地应用于迁徙飞行的遥测(Meyburg和Pfeiffer 2009年)。类似的数据,如果可以用于繁殖季节的测距行为,可以合并到基于代理的模型中,因为我们的方法足够灵活,可以合并改进的移动规则。
由于缺少可比的现场数据,我们通过两个主要的现场研究(Hötker et al. 2006, Dürr 2009)间接验证了导出的影响函数。这些研究没有考虑到死亡率与aery - wt距离的依赖性,但监测了不同地区每年每个涡轮机死亡的鸟类数量。Hötker等人(2006)估计,在德国每年约有100名红鸢被WTs杀死。假设德国的鸟撞事件分布均匀,并考虑到鸟撞事件的总数(16,500 WT;Bundesverband Windenergie (BWE) 2010),这导致每个涡轮机的年碰撞风险为0.006。第二项研究(Dürr 2009)使用了德国联邦州勃兰登堡州的记录,该地区毗邻西萨克森州。在2001年至2009年期间,红鸢与西隧相撞的次数有记录。根据这些数据,Dürr(2009)估计每个涡轮机的年碰撞率为0.028。在导出的冲击函数中,这两项研究的碰撞率分别对应2000 - 2500 m和1300 - 1500 m的aery - wt距离。这两个推导的距离范围与该地区观测到的aery - wt距离很吻合,也符合红风筝的典型飞行距离(Nachtigall 2008)。 The derived impact function is, therefore, indirectly validated.
如果在景观中不同距离处有多个小波,则与其中任何一个小波不碰撞的概率是与每个小波不碰撞的概率的乘积。各个小波的碰撞概率可以从导出的碰撞函数中得到。或者,模型可以与模型视图中的几个wt同时运行。因此,该模型也可用于评估风力发电场的效果。
碰撞风险对种群动态的影响的研究是有趣的,因为尚不清楚是WTs造成的额外死亡率导致了种群数量的下降,还是它们只是补偿了密度依赖性的种群调节。使用VORTEX种群生存能力分析软件包(参见http://www.vortex9.org/vortex.html), Hötker等人(2006)证明了由于纳入了wt引起的死亡率而导致的人口下降的明显趋势。该研究的结论是,需要更好的关于与大型交通工具相撞造成的死亡率的数据集。我们计算的死亡率和由此产生的影响函数可作为这种人口生存能力分析的投入,因此可直接有助于更好地估计人口趋势。此外,可以利用影响函数来评估禁止架设水塘的鸟笼周围缓冲区的大小,以最大限度地减少风力发电发展对鸟类的负面影响。
我们的建模方法也可以应用到其他种类的迅猛龙,只要适当调整觅食行为和参数。适应其他区域也是可能的。在这两种情况下,都需要足够的字段数据来参数化模型。特别是,比较获得的不同物种和不同景观的影响函数,从而支持对风力发电发展的生态后果的全面评估,将是有趣的。
结论
基于智能体的建模方法的分析表明,通过碰撞风险作为aerie-WT距离函数的聚合关系,可以捕捉到觅食行为和景观结构的协同效应。因此,我们通过基于代理的建模获得影响函数的方法为风力发电发展的景观规划提供了一个有用的工具,并有助于以可持续的方式实现可再生能源目标。推导出的影响函数,结合Eichhorn和Drechsler(2010)提出的生态经济分析,可以确定风能生产和红风筝保护之间的所谓权衡。这些权衡显示了增加风能生产和由此产生的生态影响之间的功能关系。
对特定景观中特定物种的影响函数进行定量说明,包括通过实地数据进行验证,对于将其用于风力发电开发的具体规划和设计过程非常重要。因此,对于濒危物种来说,必须有足够的野外数据来参数化和验证这种基于agent的模型。
致谢
我们要感谢Winfried Nachtigall和Herman H ? tker在模型开发过程中对鸟类学问题的实证支持和建议。我们还要感谢Jan Engel和Andreas Thiele,感谢他们的讨论、技术支持和编程提示。作者感谢教育和研究部资助“风能利益冲突”研究项目[Nachhaltige Landnutzung im Spannungsfeld umweltpolitisch konfligierender Zielsetzungen am Beispiel der windenergiewinnung](批准号:01UN0601A, B),作为Fona项目“可持续发展的经济学[Wirtschaftswissenschaften f & r Nachhaltigkeit] (WIN)”的一部分。这项工作得到了Helmholtz冲动和网络基金通过Helmholtz跨学科环境研究研究生院(HIGRADE)的进一步支持。
文献引用
ATKIS。2007.权威地形图信息系统ATKIS®-DGM25。许可证号码:3219/07,军械测量萨克森,德累斯顿,德国。Band, W. M. Madders和D. P. Whitfield. 2007。开发评估风电场鸟类碰撞风险的现场和分析方法。M. de Lucas, G. F. E. Janss和M. Ferrer,编辑,第259-275页。鸟类和风力发电场:风险评估和缓解。Quercus,马德里,西班牙。
巴里奥斯,L.和A.罗德里格斯,2004。岸上风力涡轮机的飞禽死亡率的行为和环境相关性。应用生态学杂志41:72 - 81。http://dx.doi.org/10.1111/j.1365-2664.2004.00876.x
鲍尔,M.范·迪瑟,k . a。Høgda, M. Klaassen, J. Madsen. 2008。气候驱动的停留地点对北极鹅迁移时间表和适应性的影响。应用生态学杂志77:654 - 660。http://dx.doi.org/10.1111/j.1365-2656.2008.01381.x
生命现象研究》2003。鸟类建模补遗Timewell和Meredith 2002。附录6。向水电公司报告。生物多样性研究,墨尔本,澳大利亚。
勃兰登堡州鸟类保护中心。2010.德国的Vogelverluste和Windenergieanlagen。Zentrale Fundkartei。勃兰登堡州:托拜厄斯Dürr。BSBCC,勃兰登堡,德国。(在线)网址:
http://www.mugv.brandenburg.de/cms/media.php/lbm1.a.2334.de/wka_vogel.xls.
Bruderer和A. Boldt, 2001。鸟类飞行特性:一、速度的雷达测量。宜必思143:178 - 204。http://dx.doi.org/10.1111/j.1474-919X.2001.tb04475.x
Bundesverband Windenergie e.v. (BWE)。2010.德国2009年的Die Entwicklung der Windenergie。(在线)网址:
http://www.wind-energie.de/de/statistiken/charts/365/0/statistik.csv.
张伯伦,dr . Rehfisch, A. D. Fox, M. Desholm和S. Anthony, 2006。规避率对风力发电机碰撞风险模型所作鸟类死亡率预测的影响。宜必思148:198 - 202。http://dx.doi.org/10.1111/j.1474-919X.2006.00507.x
迪安吉利斯,D. L.和W. M. Mooij. 2005。基于个体的生态和进化过程建模。生态学、进化与系统学年评36:147 - 168。http://dx.doi.org/10.1146/annurev.ecolsys.36.102003.152644
M. de Lucas, G. F. E. Janss, D. P. Whitfield, M. Ferrer. 2008。风电场中迅猛龙的碰撞死亡率与迅猛龙的数量无关。应用生态学杂志45:1695 - 1703。http://dx.doi.org/10.1111/j.1365-2664.2008.01549.x
德威特,A. L.和R. H. W.兰斯顿。2006。评估风力发电场对鸟类的影响。宜必思148:29-42。http://dx.doi.org/10.1111/j.1474-919X.2006.00516.x
·杜尔,t . 2009。祖尔Gefährdung des RotmilansMilvus Milvus德国的durch Windenergieanlagen。Informationsdienst Naturschutz Niedersachsen29:185 - 191。
Eichhorn, M.和M. Drechsler. 2010。农业景观中风力发电和鸟类避碰之间的空间权衡。生态和社会15: 10。(在线)网址://www.dpl-cld.com/vol15/iss2/art10/.
埃里克森,W. P., G. D.约翰逊,M. D.斯特里克兰德,D. P.杨,K. J.瑟卡,R. E.古德。2001。鸟类与风力涡轮机的碰撞:美国现有研究的总结和鸟类碰撞死亡率的比较。NWCC和RESOLVE公司,加拿大安大略省国王城,LGL有限公司和RESOLVE公司,美国华盛顿特区。
格林,V, U.伯杰,F.巴斯蒂安森,S.埃利亚森,V.吉诺特,J.吉斯克,J.戈斯-卡斯特德,T.格兰德,S.海因茨,G.豪斯,A.胡特,J.尤普森,C. J.罗根森,W. M.穆伊,B. Müller, G.佩尔,C.皮欧,S. F.莱瑞贝克,A. M.罗宾斯,M. M.罗宾斯,E.罗斯曼尼斯,N. Rüger, E.斯特兰德,S.苏伊西,R. A.斯蒂尔曼,R.瓦布ø, U.维瑟,D. L.迪安吉利斯。2006.用于描述基于个体和基于代理的模型的标准协议。生态模型198:115 - 126。http://dx.doi.org/10.1016/j.ecolmodel.2006.04.023
格林,V.和s.f. Railsback, 2005。基于个体的建模和生态。普林斯顿理论与计算生物学系列。普林斯顿大学出版社,美国新泽西州普林斯顿和英国牛津。http://dx.doi.org/10.1016/s0304 - 3800 (98) 00186 - 0
格林、V、E.雷维拉、U.伯格、F.耶尔奇、W. M.穆伊、S. F. Railsback、H. H. Thulke、J.韦纳、T.韦根和D. L. DeAngelis。2005.基于智能体的复杂系统的面向模式建模:来自生态学的教训。科学310:987 - 991。http://dx.doi.org/10.1126/science.1116681
K.-M Hotker H。Thomsen和H. Jeromin, 2006。开发可再生能源对生物多样性的影响:以鸟类和蝙蝠为例——事实、知识差距、进一步研究的需求以及发展可再生能源开发的鸟类学指南。迈克尔-奥托纳布研究所,贝尔根胡森,德国。
亨特,g . 2002。金鹰在一个危险的景观:预测缓解风力涡轮机叶片撞击死亡率的影响。顾问报告500-02-043F, PIER -环境区。准备:加州能源委员会,萨克拉门托,加利福尼亚州,美国
休斯顿,M., D. DeAngelis, W. Post, 1988。新的计算机模型统一了生态理论。生物科学38:682 - 691。http://dx.doi.org/10.2307/1310870
Jedicke, e . 2010。Rotmilan Windkraft。Naturschutz和Landschaftsplanung43:35。
约翰斯特,R.布兰德尔,R.普费弗。2001。在零散而动态的景观中觅食:人类的土地利用和白鹳。生态应用程序11:60 - 69。http://dx.doi.org/10.1890/1051 - 0761 (2001) 011 (0060: FIAPAD) 2.0.CO; 2
Krijgsveld, K. L., K. Akershoek, F. Schenk, F. Dijkf, S. Dirksen. 2009。鸟类与现代大型风力涡轮机的碰撞风险。Ardea -荷兰鸟类学家联盟的官方期刊97:357 - 366。http://dx.doi.org/10.5253/078.097.0311
Larsen, J. K.和M. Guillemette. 2007。风力涡轮机对越冬普通羽绒飞行行为的影响:栖息地使用和碰撞风险的启示。应用生态学杂志44:516 - 522。http://dx.doi.org/10.1111/j.1365-2664.2007.01303.x
J. M.勒库纳和C. Ursúa。2007.纳瓦拉(西班牙北部)风力发电厂的鸟类死亡率。177 - 192页在M. de Lucas, G. Janss, M. Ferrer. 2007。鸟类和风力发电场:风险评估和缓解。Quercus/利比里亚Linneo,马德里,西班牙。
M. Madders和D. P. Whitfield. 2006。高地猛禽和风电场影响的评估。宜必思148:43-56。http://dx.doi.org/10.1111/j.1474-919X.2006.00506.x
定,2010年。Aktueller Wissensstand zum Konfliktfeld“Rotmilan和Windkraft”。(在线)网址:http://www.wind-energie.de/fileadmin/dokumente/Themen_A-Z/Vogelschutz/07_Mammen_Rotmilan.pdf.
Mammen, U.和T. Dürr。2006.Rotmilane und Windkraftanlagen - Konflikt oder übertreibung?APUS - Beiträge zur aviflora Sachsen-Anhalts13:73 - 74。
麦格雷迪,M. J., D. R. A.麦克劳德,S. J.佩蒂,J. R.格兰特,i.p.班布里奇。1997。金雕和林业。林业委员会研究资料说明292。英国罗斯林林业委员会。
麦克劳德,D. R. A, D. P.惠特菲尔德,A. H.菲尔丁,P. F.霍沃斯,M. J.麦克格雷迪。2003.预测金鹰的本垒打范围Aquila chrysaetos在苏格兰西部。鸟类科学2:183 - 198。
梅伯格,B. U.和T. Pfeiffer. 2009。gps -卫星-遥测als neue Methode, der Forschung am RotmilanMilvus Milvus.Informationsdienst Naturschutz Niedersachsen29:211。
Nachtigall, w . 2008。Der Rotmilan (Milvus Milvus, L 1758)在萨克森和Südbrandenburg - Untersuchungen zur Verbreitung和ökologie。论文,Martin-Luther-Universität Halle - Wittenberg。(在线)
URL:http://sundoc.bibliothek.uni-halle.de/diss-online/08/08H218/prom.pdf.
欧尔,C.和M.埃希霍恩,2010。德国风电发展的区域空间规划与国家上网电价合格标准之间的不匹配——西萨克森州的一个案例研究。土地使用政策27:243 - 254。http://dx.doi.org/10.1016/j.landusepol.2009.06.004
皮尔斯-希金斯,J. W., L.斯蒂芬,R. H. W.兰斯顿,i.p.班布里奇,R.布尔曼。2009。高地风电场周围繁殖鸟类的分布。应用生态学杂志46:1323 - 1331。
Percival, s.m.2000。英国的鸟类和风力涡轮机。英国野生动物12:8-15。
珀西瓦尔,S. M. 2005。鸟类和风力发电场:真正的问题是什么?英国鸟类98:194 - 204。
Podolsky, r . 2003。鸟类碰撞风险(ARC)模型。NWCC生物学意义研讨会,2003年11月17-18日。国家风协调委员会,华盛顿特区,美国。
Podolsky, r . 2005。风险评估工具的应用:鸟类碰撞风险模型。86 - 87页在s·s·施瓦茨,编辑。陆地野生动物与风的相互作用,研究会议第五,弗吉尼亚州兰斯顿,2004年11月3日至4日。为国家风协调委员会野生动物小组委员会准备。RESOLVE, Incorporated, Washington, d.c., USA。
Rodríguez, C., K. Johst和J. Bustamante. 2006。作物类型如何通过猎物质量和可得性影响小红隼的繁殖成功?建模方法。应用生态学杂志43:587 - 597。http://dx.doi.org/10.1111/j.1365-2664.2006.01152.x
苏格兰国家遗产(SNH)。2000.风电场和鸟类:计算理论碰撞风险假设没有避免行动。指南系列,苏格兰自然遗产。(在线)网址:http://www.snh.gov.uk/docs/C205425.pdf.
苏格兰国家遗产(SNH)。2010.碰撞风险评估——Excel计算模板.(在线)的URL: http://www.snh.gov.uk/docs/C234672.xls。
斯梅尔斯和S.缪尔,2005。模拟了风电场对塔斯马尼亚楔尾鹰在整个物种范围内的累积影响。项目没有。4857.向环境及文物局提交报告。生命现象研究企业。有限公司
斯莫尔伍德,K. S.和C.西兰德,2007。加州Altamont风资源区的鸟类死亡率。野生动物管理杂志72:215 - 223。http://dx.doi.org/10.2193/2007-032
塔克,1996年。鸟类与风力涡轮机转子碰撞的数学模型。太阳能工程学报118:253 - 262。http://dx.doi.org/10.1115/1.2871788
Verboom, J., A. Schotman, P. Opdam和J. A. J. Metz. 1991。欧洲五子兽在破碎农业景观中的元种群。Oikos61:149 - 156。http://dx.doi.org/10.2307/3545332
惠特菲尔德博士,2009。在“带状”碰撞风险模型下的风电场金鹰避碰研究。自然研究有限公司,班奇利,英国。
魏菲尔德,M.马得斯,2006a。风电场对鸡鹞影响的综述马戏团cyaneus以及对避碰率的估计。自然研究信息注1,自然研究有限公司,班奇利,英国。
D. P.威特菲尔德和M.马得斯。2006b。推导红鸢的避碰率Milvus Milvus。自然研究信息笔记。自然研究有限公司,班奇利,英国。
Wichmann, m.c., W. R. J. Dean和F. Jeltsch. 2004。全球变化对褐雕的挑战(Aquila rapax):就预测的气候和土地用途变化,模拟灭绝风险。鸵鸟75:204 - 210。http://dx.doi.org/10.2989/00306520409485446
韦根,T., F. Jeltsch, I. Hanski和V. Grimm, 2003。利用模式导向建模揭示隐藏信息:协调生态学理论与应用的关键。Oikos100:209 - 222。http://dx.doi.org/10.1034/j.1600-0706.2003.12027.x
威廉姆斯,P. H., D.吉本斯,C.马勒斯,A.雷贝洛,C.汉弗莱斯和R.普莱斯西。1996。英国鸟类多样性保护的丰富度热点、稀有度热点和互补区比较。保护生物学上午10 - 74。http://dx.doi.org/10.1046/j.1523-1739.1996.10010155.x
