|
版权所有©弹性联盟1998*
圣克莱尔,C. C., M. Bélisle, A. Desrochers, S. Hannon, 1998。森林鸟类对栖息地廊道和林隙的冬季响应。保护生态(在线)2(2): 13。可从互联网上获得。URL: http://www.consecol.org/vol2/iss2/art13/
在本文的一个版本中,文本、图形、表格和附录是分开的文件,可以通过下面的这个文件找到链接.
研究 森林鸟类冬季对栖息地廊道和林隙的响应科琳·卡萨迪·圣克莱尔1、2,Marc Belisle2,安德烈Desrochers2,苏珊Hannon1
1阿尔伯塔大学;2大学�拉瓦尔
森林的破碎化和栖息地的丧失可能会干扰森林栖息鸟类的迁徙或扩散。尽管人们对这些影响的严重性和缓解它们的方法很感兴趣,但对不同栖息地类型的实际移动模式知之甚少。我们研究了过冬的留鸟在蜂群叫声的吸引下的活动,以比较森林鸟类在连续的森林中,沿着狭窄的走廊(围栏),以及在森林覆盖的缝隙中穿越不同距离的意愿。我们也量化了黑顶山雀的意愿(Poecile atricapillus),以便在有其他可供选择的森林绕道路线时穿越林间。当距离增加到200米时,所有的物种都不太可能对这种叫声做出反应,尽管白胸斑胸草(Sitta有害无益)及毛啄木鸟(Picoides绒毛)通常比山雀和绒啄木鸟(p .下毛竹).山雀在走廊和连续的森林中行走的可能性一样大,但随着林隙距离的增加,它们不太可能穿过林隙。相对于森林,其他物种不太愿意在廊道和林隙中活动,生境间的差异也随着距离的增加而增大。对于山雀来说,在有森林绕道的情况下,它们的选择在我们测试的距离范围内不同,主要受绕道效率的影响(相对于可用绕道的捷径长度)。在短距离飞行中,鸟类会选择森林里的迂回路线,不管它们的效率如何。随着绝对距离的增加,当绕行效率较低或初始距离较高时,鸟类倾向于在开阔地带使用较大的捷径,但它们将与最近森林边缘的距离限制在25米。因此,山雀在有森林替代选择时不愿意跨越50米的林隙,但在没有替代选择时,它们有时会跨越200米的林隙。我们的结果表明,走廊的存在增强了一些(但不是全部)森林鸟类的活动,甚至对林隙宽度不太敏感的山雀也不愿冒险远离森林覆盖层。
关键词:黑顶山雀,走廊,绕路试验,扩散,绒毛啄木鸟,森林破碎,缝隙宽度,毛啄木鸟,运动,Poecile atricapillus,Picoides下毛竹,Picoides绒毛,Sitta有害无益白胸五子雀。
在世界各地的森林地区,栖息地碎片化现象日益严重,并对森林鸟类产生了各种负面影响(参见Wilcove等人1986年、Saunders等人1991年、Opdam等人1994年、Robinson等人1995年的评论)。除了栖息地丧失的相关和更严重的影响(McGarigal和McComb 1995, Fahrig 1997)外,碎片化还通过阻碍鸟类在剩余栖息地斑块之间的移动,改变了它们的种群结构(Lens和Dhondt 1994, Matthysen等人1995,Matthysen和Currie1996)。不幸的是,很难量化限制个体移动的栖息地碎片化的水平和结构,特别是对鸟类这样的流浪物种。到目前为止,关于栖息地碎片化的新兴文献中,大多数重点关注的是栖息地斑块的占用和丰度模式,其分散机制在很大程度上被假定或忽略(Wiens等人1993年,Ims 1995年)。在行为生态学中,长期以来被认为影响动物运动决策的因素(例如,学习、捕食风险、旅行成本)在元种群和景观生态学中仍然没有对应的因素(另见Wiens等人1993年,Lima和Zollner 1996年)。
由于从定义上讲,元种群是由个体的移动联系在一起的(Gilpin and Hanski 1991, Opdam 1991),因此对栖息地斑块之间生物移动的详细了解(即通过矩阵)对于预测碎片化对种群的影响至关重要。矩阵阻碍运动的程度在很大程度上取决于它的空间构型(Taylor et al. 1993, Fahrig and Merriam 1994, Ims1995),反过来,动物在不同和高度特定的空间尺度上感知矩阵(Kotliar and Wiens1990, Keitt et al. 1997)。在粗略的尺度上,某一特定的动物只能借助斑块之间的物理连接或走廊来绕过不适宜居住的基质。这些走廊已经被证明间接地增加了栖息地斑块中不同物种的活动(例如,Wegner和Merriam 1979, Dunning等人1995,Haas 1995, Machtans等人1996),尽管关于它们的效用有很多争论(参见Simberloff和Cox 1987, Saunders和Hobbs 1991, hobs1992, Simberloff等人1992,Beier和Noss 1998的评论)。在更精细的尺度上,个体仍然可以在矩阵中感知到足够的连通性,从而允许补丁之间的移动(Wiens 1994, 1995, Ims 1995),这也可以被描述为感知范围(Lima and Zollner 1996, Zollner and Lima 1997)或间隙交叉能力(Desrochers and Hannon 1997)。动物是否会穿越斑块之间的间隙,可能会因物种、斑块栖息地类型、基质类型以及许多通常与景观生态无关的因素而有所不同,如天气变量、季节、替代路线的存在以及相关成本(如感知到的捕食风险、绕行长度)。
要量化森林物种在走廊和林隙中的活动一直很困难,部分原因是调查人员经常既不能确定动物在林隙间活动的原因,也不能提供动物在林隙间活动的动机。鸟类鸣叫的录音回放为鸟类研究提供了这种动力。例如,特定物种的领地歌声被用来量化森林鸟类进入不同矩阵类型的意愿(Sieving et al. 1996)以及穿越毗邻领地100米的开放区域的间隙(Rail et al. 1997)。Desrochers和Hannon(1997)利用黑顶山雀(Black-capped Chickadees)的围捕叫声(Poecile atricapillus),以研究几种森林物种的繁殖后扩散。通过诱导鸟类穿过或绕道森林覆盖的林隙,他们发现大多数物种对50米窄的林隙表现出抵抗力,而且鸟类通常更喜欢林地路径,即使这些路径比它们的林隙长三倍。虽然三项研究都表明,生境缺口对森林鸟类造成的问题可以通过走廊加以缓解,但仍需将走廊与基质利用进行直接比较(Nicholls和Margules1991, Inglis和Underwood 1992, Simberloff等人1992)。还需要更好地解决决定鸟类是否会穿过缝隙的因素。这些因素可能包括生境边缘的配置、空间尺度和其他个体的影响。
在这里,我们结合了两个数据集来解决这些需求,通过描述在三种栖息地类型(连续的森林、间隙和走廊)中过冬的留鸟对回放的反应。两项研究都假设森林鸟类在开阔的区域会有更高的被捕食风险,而且两项研究都将重点放在冬季的常住物种上,原因有三:常住物种可能对栖息地破碎化的影响更敏感(Bierregaard et al. 1992, Wiens 1994);当环境和食物压力可能最大时,冬季可能会约束行为(Ekman 1984, Desrochers et al. 1988, McNamara和Houston 1990);冬季场地逼真度的影响可能会进一步限制居民的迁移。我们解决了以下问题:(1)当栖息地是连续的森林、森林走廊或森林斑块之间的间隙时,鸟类对包围叫声的反应意愿有何不同?(2)这些反应在物种间有何不同?(3)当鸟类可以选择完全在森林覆盖下飞行时,哪些因素会影响它们穿越的间隙的大小?
我们研究了1996年1月10日至1997年3月9日期间森林鸟类在两个以农业为主的地区的活动模式,一个在阿尔伯塔省中北部北方针叶林的南端(北纬54°37′,西经113°20′),一个在Québec市附近的圣劳伦斯低地的混合森林遗迹(北纬46°45′,西经71°20′)。这两个地区都有大约20%的森林覆盖,其余部分用于种植作物、放牧和住房。在艾伯塔省,这些森林主要以杨树(美洲山杨而且p . balsamifera)和白云杉(云杉glauca);在Québec,由sugar maple (宏碁蔗糖)、黄桦(桦木属alleghaniensis)、山毛榉(水青冈属grandifolia)、香脂冷杉(冷杉属balsamea).
在这两个地方,我们都播放了黑顶山雀和红胸五子雀(Sitta黄花),以吸引冬季森林鸟类前往目的地。在几种森林类型和季节中,山雀的围攻叫声似乎对吸引不同物种非常有效(Hurd 1996, Desrochers和Hannon 1997)。为了便于研究之间的比较,我们使用了相同的录音,并遵循了Desrochers和Hannon(1997)中相同的广播方法。这些呼叫通过一个连接在便携式磁带播放机上的5瓦扬声器播放,人类听众可以从大约225米远的地方听到。研究人员认为,鸟类在距离广播喇叭水平10米以内飞行时,会对播放的声音做出反应。我们没有在阵风或中到大风、温度低于-30°C或中到强降水期间进行试验。
栖息地的试验在艾伯塔省,我们进行了栖息地试验,我们比较了三种栖息地类型的鸟类的旅行意愿:连续的森林;走廊<10 m宽的以杨树为主的森林(围栏);而且差距在森林覆盖斑块之间(图1).有关该研究区域的更详细描述,请参阅Hannon和Cotterill(1998)。在每次试验中,我们在一个指定的地点播放围攻叫声最多6分钟,或者直到至少有两只鸟到达而没有新的鸟到达为止1分钟。原始磁带的目的是为试验提供一个已知的起点。当最初的带子停止时,另一个观察者在目的地开始播放磁带,播放6分钟或直到所有的起源鸟都做出反应。两名观察者都观察了鸟类的飞行路线,以确定对目的地带子做出反应的鸟类的数量和种类。当一个或多个来自起源的个体做出反应时(在山雀群、五子雀或啄木鸟对中,反应通常是相同的),我们为给定物种的试验打分。当一个物种在起源处没有被检测到的个体也出现在终点时,反应得分为阴性。每次试验,我们也记录时间、温度(+5°C),以及云量(阴天、偏云、晴天)、风速(无风、微风、稳风)和降水(无风、小雨或雪)的相对测量(2 - 3点尺度)。出发地与目的地之间的距离在1:20 000航拍地图(间隙)上测量,或通过预校准步距测量,精度为5米。试验在3种生境类型中随机分配,在20 ~ 200 m距离范围内近似平均分配。试验在9点到16点之间进行。为避免习惯效应,所有试验在空间上间隔至少400米或在时间上间隔至少2周。廊道的起源地和林隙的起源地和目的地都由面积至少为0.5 ha的森林斑块组成。
图1所示。在三种生境类型中指示起源(圆圈)和目的地(三角形)扬声器位置的生境试验示意图表示。试验距离从25米到200米不等。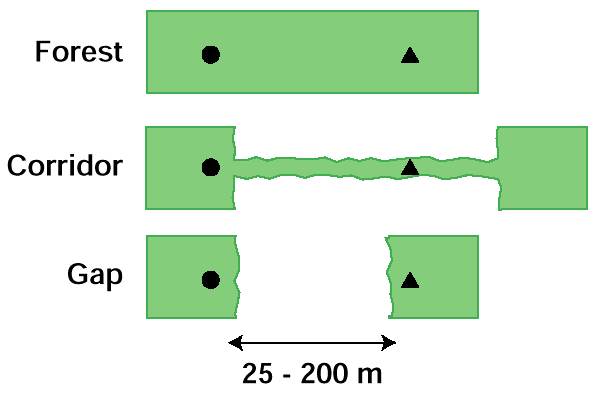
|
绕道试验
我们进行了绕道在Québec上进行的试验,旨在提供鸟类在森林覆盖中出现缺口时做出的决策规则的更精确的分辨率。在这里,我们重点研究了黑顶山雀,并使用了森林边缘结构,在这种结构中,鸟类必须在通过缺口(即捷径)直接到达目的地之间进行选择,或在森林覆盖下选择更长的路线(图2;参见Desrochers和Hannon, 1997)。在目标地点只播放了10分钟的围攻叫声,而两名观察员观察了在森林边缘10米范围内出现的鸟类。我们没有把鸟类吸引到预先确定的起源,因为当它们到达森林边缘时很容易被看到。在回放过程中,视觉上尽可能多地跟踪鸟类的路径,直到它们到达目的地或试验结束。使用视觉地标和激光测距仪(精度1米),我们随后为每个有反应的个体绘制了这些路径。为了限制交叉决策的干扰,在整个试验过程中,观察者与起点边缘和终点的距离至少为30米。目标地点和最近的原始森林边缘之间的距离是随机选择的,但大约是有规律的,在10到200米之间。通过改变这些距离,我们创造了在开放的初始距离的变化,以及相应的绕行长度的变化(图2).试验在8点到16点之间进行。试验点仅使用一次,且间隔至少500米。
图2所示。表示典型森林边缘配置的迂回试验的示意图表示。三角形表示说话者的位置,圆圈表示第一眼看到的焦点鸟(#1和2)的位置。对于每只焦点鸟,我们记录了它在开阔环境中到达扬声器的最短距离(DO我)、森林覆盖下可用路线的距离,以及存在同种的数量。记录鸟类在野外的实际活动路径(DO一个)和森林覆盖下(DF一个).注意,试验在u型(如鸟1号)和v型(鸟2号)两种情况下进行。因变量包括发生在露天的实际路径的比例(ε一个/(ε一个+εDF一个)),危险区域定义在鸟的路径和相邻的森林之间,和森林边缘的距离(美联储)描述山雀在给定的捷径上冒险到森林边缘的最大距离。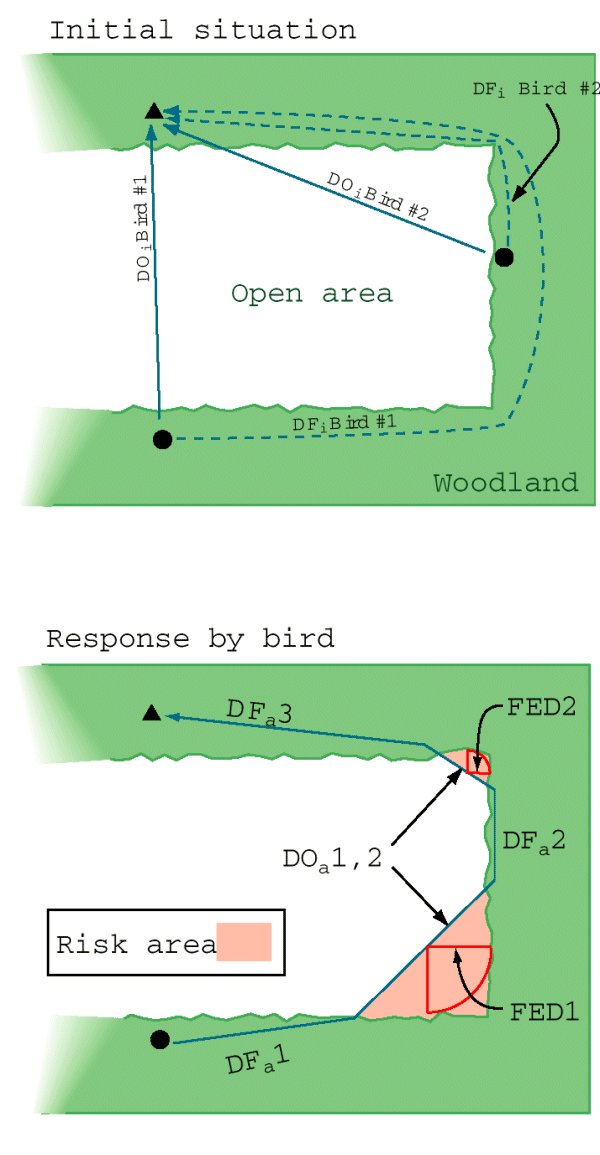
|
森林边缘形态的变化(图2)生成了三个独立变量,用于后续的分析:在开放空间中,鸟类在起源边缘的探测点与目的地之间的初始距离(DO我,为比例因子);开阔地的初始距离与通过相邻森林的迂回路线的长度之比(DO我/ DF我,绕行比);以及这两项之间的相互作用(DO我x做我/ DF我).为简单起见,我们考虑绕行比(DO我/ DF我)是绕行效率的指标。因此,绕行比的低值对应较长的绕行长度相对于开放的初始距离(有效的捷径),而高值表明两条路线的长度相似(低效的捷径)。最后,我们记录了同族群体大小(在试验中观察到的个体数量)作为一个额外的自变量。我们使用三个连续的因变量作为合理的衡量标准,来衡量鸟类在森林中昂贵(在时间和精力方面)但更安全的路线与在开阔地带的捷径之间做出的权衡。在开阔地带,较低的旅行成本可能伴随着更大的捕食风险。(1)比例这只鸟在野外走的路的长度表明它相对使用了一条或多条捷径。(2)尺寸危险区域这只鸟从邻近森林的路径发散形成的路径,整合了两个重要的衡量标准,即路径在开阔地带的长度和路径到森林边缘的距离。(3)鸟冒险离开森林边缘的最大距离(森林边缘的距离)测量它到安全的感知距离。
统计分析我们使用逐步逻辑回归和反向消除来衡量自变量在生境试验中的影响。这些模型中变量的进入和去除阈值分别设为alpha = 0.05和alpha = 0.10。我们使用Wald统计量和参考类别来检验生境类型和物种类别变量之间多重比较的显著性,并报告R值,这类似于线性回归中的偏回归系数,以描述这些关系的方向和强度(Norusis 1994)。因为我们预计响应的概率会随着距离的增加而降低,而且在栖息地试验中,走廊相对于缝隙和森林相对于走廊的响应概率会更大,所以我们报告了这些变量的单侧概率。对于迂回试验的连续因变量,当方差为异质性时,我们采用多元线性回归并进行变量变换。一些自变量以其降低共线性的方法为中心(Kleinbaum et al. 1988)。绕行试验中的混淆尺度效应可能源于DO之间的相关性我/ DF我比和做我(r年代= 0.13,P= 0.09,n=158),在控制了其他自变量的贡献(第三类平方和)后,回归评估每个自变量的贡献。
同种吸引力的潜在影响会限制群体成员之间行动决策的独立性(Smith和Peacock 1990, Reed和Dobson 1993, Romey 1996),通过两种方式来解决。在栖息地试验中,如果一个或多个个体出现在目的地,每个相同的鸟群产生一个响应,那么每个物种被认为是积极响应。这无疑高估了积极的反应,但避免了栖息地类型与个体检测的容易程度相混淆,这在森林试验中是比较困难的。在绕行试验中,每只鸟的路径被单独记录,并被赋予一个统计权重,该权重等于在同一试验中记录的路径数的倒数(即,每一次试验被赋予一个自由度,而构成该试验的鸟的路径被分析为一个自由度的一部分)。
在栖息地试验中,鸟类成功地被吸引到起源处92次,涉及11个物种的408只个体。除2项试验外,其余试验均为Black-capped Chickadees,且大多数(61%)试验仅限于该物种。在7次或更多的试验中,只有3种其他物种出现在了随后的分析中:Sitta有害无益,n= 10)、啄木鸟(Picoides下毛竹,n= 8)、毛啄木鸟(p .绒毛,n= 7)。我们用逻辑回归分析了距离、栖息地类型、距离与栖息地类型的相互作用以及物种(n= 115)。反应概率随距离的增加而降低(卡方= 8.0,df = 1,P= 0.003),且受两种植物影响均显著(χ 2 = 16.6, df = 3,P= 0.0009),栖息地类型与距离的交互作用(卡方= 19.5,df = 2,P< 0.0001)。对这种相互作用的进一步研究表明,随着距离的增加,鸟类在林隙中的反应要比在森林中的低(Wald = 13.8,R= -0.30, df = 1,P= 0.0001),但走廊和森林的响应没有区别(Wald = 0.6,R= 0.0, df = 1,P= 0.2)。与山雀相比,五子雀和毛啄木鸟对回放的反应更弱(Wald = 12.1,R= -0.27, df = 1,P= 0.0005;瓦尔德= 3.7,R= -0.11, df = 1,P= 0.06,但Downy woodpecker与山雀没有区别(Wald = 0.5, df = 1,R= 0.00,P= 0.5)。没有一个时间或天气变量达到进入模型的标准。由于整个样本受到山雀反应的严重偏差,我们测试了距离、栖息地类型和距离x栖息地类型相互作用对五子雀和啄木鸟的影响(n= 25)。距离再次对反应产生负面影响(卡方= 4.8,df = 1,P= 0.03)和交互项(卡方= 8.4,df = 2,P= 0.01)。与森林相比,林隙的响应可能性较小(Wald = 4.1,R= -0.25, df = 1,P= 0.02)和走廊(Wald = 3.2,R= -0.19, df = 1,P= 0.04),但走廊和缝隙没有差异(Wald = 0.2,R= 0.0, df = 1,P= 0.5)。
我们只对山雀重复了这个分析(n= 90)得出他们反应的预测模型。同样,反应概率随距离的增加而降低(卡方= 6.0,df = 1,P= 0.01),距离与生境类型的交互作用(卡方= 13.8,df = 2,P= 0.001)。对于第一个模型,与林隙相比,森林的距离x栖息地类型相互作用较强(Wald = 9.3,R= 0.29, df = 1,P= 0.002),但与走廊相比,森林则不然(瓦尔德= 0.02,R= 0.0, df = 1,P= 0.5)。该模型的预测值表明,山雀在森林中穿行200米的可能性是在走廊中穿行200米的4倍,在走廊中穿行200米的可能性是在走廊中穿行的3.5倍。图3).
图3所示。黑顶山雀对山雀在森林、走廊和林隙栖息地距离增加时发出的围捕声作出反应的概率。预测概率由逻辑回归(n= 90)。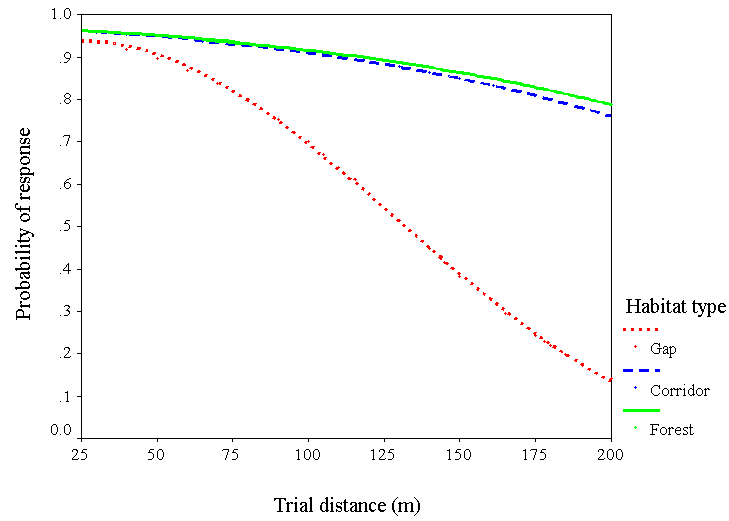
|
绕道试验
我们进行了151次试验,描述了297条山雀个体的路径。森林边缘配置主要通过迂回比(DO)影响山雀的路径选择我/ DF我),它们占了所有三个连续响应变量(表1).山雀在开放环境下的路径长度的比例[EpsilonDO]一个/ (EpsilonDO一个+ EpsilonDF一个随DO增加我随绕行比的增加而减小(表1).然而,绕行比的影响不是恒定的,这表明两个自变量之间的显著负交互作用(DO我/ DF我,做我;表1).这意味着,当山雀倾向于不考虑路径长度而选择绕行路径时,绕行效率会影响距离较远的户外路径长度的比例,但在距离较短的地方影响不大。例如,当绕行比超过0.7和DO时我当< 115 m时,迂回比的预测价值不大,山雀在野外的路径长度所占的比例变化较大,在0 ~ 1之间。相比之下,绕行比是非常重要的,当DO我>为150 m,捷径长度随捷径比从0.85降低到0.75而增加最急剧。风险区域的大小(s)和森林边缘距离(s)给出了类似的结果(表1),也表明当绕行比降低、开放初始距离增加时,山雀会选择较长的捷径。自变量之间的负相互作用也许可以用森林边缘距离的响应变量来最好地说明;绕行比对山雀从森林边缘冒险的距离有较大的影响我增加(图4).因此,当山雀在开阔的森林中面对较大的初始距离和相对低效的替代路线时,它们会选择最大的捷径。
图4所示。绕行比(DO我/ DF我)和开阔地初始距离(DO我)在森林边缘距离,山雀冒险从森林边缘的距离。较低的绕行率代表在森林覆盖下绕行的效率较低。采用多元线性回归(n= 151)。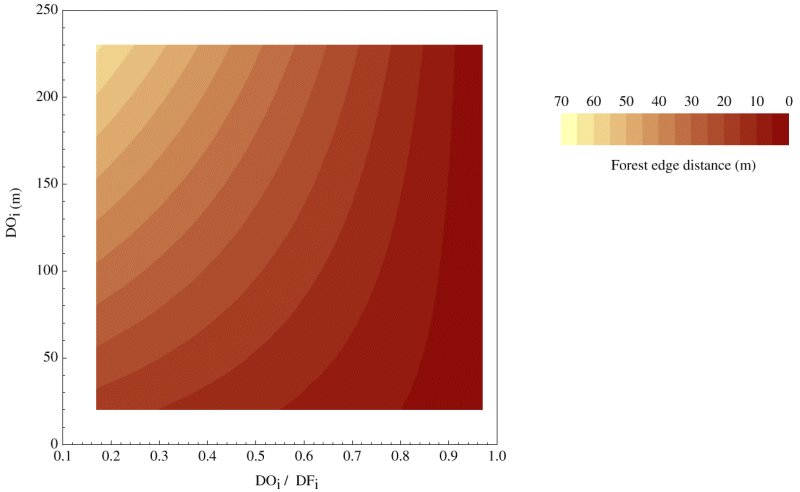
|
表1。多重线性回归(III型平方和)研究了森林边缘配置和同类群体大小对三个响应变量的影响,这些响应变量描述了黑顶山雀在迂回试验中所走路径的各个方面(n= 151;变量定义见图2)。
一个做我(80.4+33.3米,意味着+1 SD)和DO我/ DF我(0.67+0.17)以降低共线性的方法为中心。组大小平均7.3+3.3个体,1 ~ 18个体。 b为了避免在分析中混淆多个响应变量,响应变量实际上是Epsilon DO一个模型包含了总路径长度(Epsilon DO)一个+εDF一个)作为协变量。由于协变量的影响在分析中保持不变,该模型可以解释为研究其他自变量对在开放中旅行的路径长度比例的影响。 |
森林边缘距离的重要性进一步得到了一个约25米的阈值距离的说明,山雀不会冒险超过这个阈值距离(图5A和B;95%的观测路径包括森林边缘距离小于或等于24米)。这个阈值相当于50米的间隙大小,不是场地配置的人工影响;森林边缘的实际距离变化很大,但通常比鸟类沿着DO定义的路径在开阔地带穿越所能到达的最大距离要小我(图6).森林边缘距离与危险区之间的关系进一步强调了一个阈值,因为山雀愿意在u型站点中定义相对较大的危险区,只要森林边缘距离不超过25 m (图6 b).响应变量均不受同质组大小的影响(表1).
图5所示。黑头山雀冒险到森林边缘的最大距离(森林边缘距离)与初始森林绕道选择DO的效率的函数我/ DF我, (B)开口处初始距离DO的长度我.314个数据点来自151个独立的绕行试验。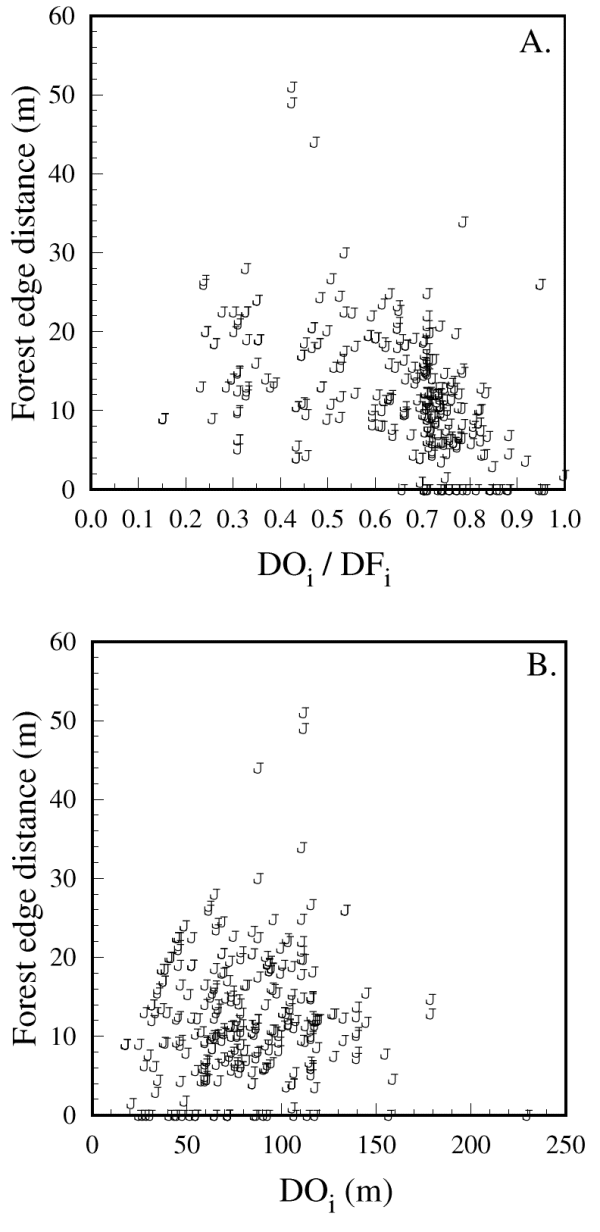
|
图6所示。黑顶山雀表现出的森林边缘距离以(a)的函数表示,如果它们沿着一条由初始距离DO定义的路径在开阔地带穿越,那么它们可能到达的森林边缘的最大距离我(B)鸟类的路径从相邻的森林边缘发散形成的危险区域。(A)中的关系斜率显著小于1 (H0:小于等于1,t= -13.3, df = 149,P< 0.0001), 95%的实际值在24 m以下(实线)。314个数据点来自151个独立的绕行试验。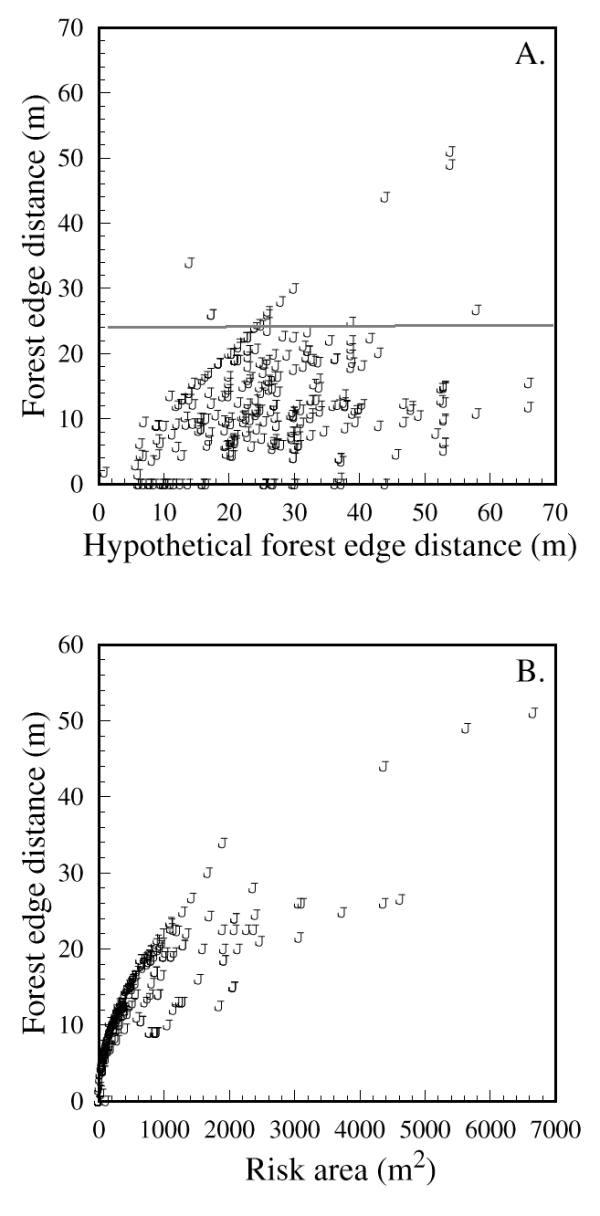
|
结果表明,像简单的栅栏那样狭窄的廊道确实促进了一些冬季鸟类的活动,而且这些廊道与连续的森林覆盖同样有效地促进了山雀的短距离(< 200 m)活动。相反,栖息地间隙开始在相对较短的距离上阻碍斑块之间的移动;到100米远的地方,山雀穿越树篱的可能性比它们在森林或走廊中响应围拢的呼叫的可能性要低25%,而在200米远的地方,这种可能性降低了80% (图3).山雀无疑会穿过这么大的空隙(个人观察),但我们的结果量化了他们这样做的意愿。
尽管所有物种都不太愿意跨越栖息地间隙,但我们研究的4种冬季居民的反应不同。山雀和绒毛啄木鸟通常比白胸五子雀和毛啄木鸟表现出更积极的反应,尽管这四种鸟类有相似的生活史和栖息地特征,而且经常被观察到一起旅行。所有鸟类对空白的反应都是负面的,但五子雀和啄木鸟对走廊的反应与森林试验相似。对于这些物种来说,走廊宽度可能是至关重要的(例如,Tischendorf和Wissel 1997),而栅栏本身可能太窄,无法提供足够的覆盖。走廊使用的物种差异也可能源于不同的家庭范围大小或群集行为倾向,这两种倾向对山雀来说都更大。这种物种差异对这些物种处理栖息地碎片化的方式有重要影响。我们的发现强化了这样一个论断,即走廊使用的量化结果不能轻易地在物种或栖息地中推广(Lynch and Saunders 1991, Lindenmayer 1994)。Roland和Taylor(1997)发现,在拟寄生物群落中,对栖息地破碎化的反应可能发生在不同的空间尺度上,部分取决于体型大小。其他空间、生态和行为属性也可能影响物种对廊道的依赖。我们在一个物种——黑顶山雀(Black-capped Chickadee)的背景下研究了这些属性,目的是为量化和比较物种的反应提供一个基线。
山雀对绕行的反应分析表明,森林边缘结构是破碎化景观的一个重要属性。在所有情况下,绕行效率解释了响应变量的最大差异,但我们的结果也表明,响应可能依赖于规模。更具体地说,我们发现鸟类在野外飞行的距离通常随着绕路效率的降低而增加(即绕路率的降低),并且随着在野外飞行的初始距离而增加。这些结果表明,响应度存在线性成分,但当绕路经济且开放距离< 115 m时,响应度实际上变化很大。鸟类显然对绕行效率和绝对距离有反应,但它们对两者的敏感度似乎随着绝对距离的变化而变化。然而,许多其他变量也必须影响他们关于路径选择的决策规则。诸如胆量、优势度、同种群体大小和内部状态等因素已被证明会影响森林鸟类觅食的距离,作为觅食者对捕食风险敏感性的指标(例如,Lima和Dill 1990, Todd和Cowie 1990, Slotow和Paxinos1997)。考虑到这些影响和环境条件的潜在重要性(McNamara和Houston 1990),我们对结果中始终没有同类群体规模和天气影响感到惊讶。
风险区域和森林边缘距离的响应变量进一步强调了空间尺度的重要性,开放初始距离与绕行效率之间的相互作用再次明显。两种响应变量均随绕行效率(即较高的绕行率)而减小,随开路初始距离而增大。因此,山雀从森林边缘冒险的最大距离(森林边缘距离)仅在较大的初始距离上取决于绕行效率,而鸟类通常选择森林路径,而不考虑它在较小距离上的效率。这些结果表明,只有当达到一定的间隙阈值时,绕行选择才会成为路径选择的重要决定因素。山雀在短距离不加区分地绕行的事实表明,它们更喜欢在森林覆盖下旅行,除非它们的选择在经济上或空间上受到限制。
也许我们对绕行路线的分析中最有趣的结果是,从森林边缘明显出现了一个阈值距离,鸟类不会冒险超过这个阈值距离。这个阈值出现在25米左右,在森林边缘距离变量的所有处理中都很明显,它不是由试验开始时鸟类可以冒险到森林边缘的潜在距离的虚假限制造成的。因此,山雀在选择在森林覆盖下绕行时,似乎不愿意穿过>米的林隙(即距离边缘25米时的林隙大小),这一数值与Desrochers和Hannon(1997)在夏末的研究结果一致。其他物种也检测到了类似的间隙阈值(见Andreassen et al. 1996, Zollner and Lima 1997)。将我们的森林边缘距离阈值效应与更大的风险区域变化进行比较,可以提供一些关于山雀感知捕食风险的方式的信息。因为鸟类更容易受到距离的影响,而不是捕食者可能出现的面积的增加,所以它们可能会将任何开放区域的捕食风险相对恒定,并仅依靠靠近森林来减轻其影响。在小的空间尺度上避免缝隙进一步表明山雀对开放区域有绝对的风险。
感知范围(Lima and Zollner 1996)决定了动物是否有可能感知到一个裂口另一边的风景,可能会因季节和区域的不同而有所不同。在冬季,当鸟类不再需要保卫领地或喂养幼鸟时,这种距离可能会增加,或者随着鼓励探索的短暂食物来源(如果实)的消失,这种距离可能会减少(Lynch and Saunders 1991)。生境破碎化程度的区域差异可能会通过习惯化甚至进化变化影响感知范围。没有绕路选择的黑顶山雀在Québec在夏末几乎从来没有穿过100米大的间隙(Desrochers和Hannon 1997),但在冬季阿尔伯塔省穿过如此大的间隙的概率仅减少了25%。这种差异可能来自于在冬季跨越鸿沟的更大意愿,来自于影响碎片化反应的区域差异,或来自于成功杂交的分类差异(见方法)就会高估阿尔伯塔省的交叉倾向。
我们的研究结果对破碎化景观中森林鸟类的管理具有多种意义。首先,我们发现了黑顶山雀在相对较短的距离(200米)内愿意在走廊(围栏)和连续森林中旅行的直接证据。相比之下,栅栏似乎并没有促进五子雀和啄木鸟的移动,尽管走廊宽度的重要性应该在它们对这些物种的重要性被忽视之前进行调查。随着林隙宽度的增加,4种植物的跨林窗意愿急剧下降。显然,森林覆盖面积的巨大缺口会对冬季森林鸟类的活动产生负面影响,但对某些物种来说,这种影响可能会通过维持或创建栖息地走廊得到改善。当山雀可以选择在森林中绕行时,它们会选择在开阔的地方走捷径,这表明它们明显更喜欢在森林覆盖下行走。当绕行相对昂贵且初始开放距离较大时,山雀使用捷径,但避免50米的空隙。因此,山雀在距离森林覆盖层的绝对距离既不小也不超过明显阈值的中间条件下最有可能使用捷径。其他的居民,显然比山雀对空隙更敏感,预计会表现得更保守。在已经存在许多较大差距的地区(例如,农田),建立森林垫脚石(sensu Simberloff et al. 1992)可能会促进移动。 The usefulness of this technique merits exploration.
欢迎对本文作出回应。如果被接受发表,您的回复将被超链接到文章。要提交评论,请关注这个链接.要阅读已经接受的评论,请关注这个链接.
阿尔伯塔体育、娱乐、公园和野生动物基金会、生物多样性资助计划(由阿尔伯塔猎人和渔夫的许可费支持)、加拿大环极研究所和副院长办公室、阿尔伯塔大学、米努克生物研究站提供资金和后勤支持,博士后奖学金来自伊扎克·沃尔顿·基拉姆纪念基金(阿尔伯塔大学)和生物研究中心Forestière,Université Laval (C.C. St. Clair),获得NSERC研究生奖学金和PQSPB凯利基金研究资助(M. Bélisle),以及NSERC操作资助(a . desrochers和S. Hannon)。我们很感激那些允许我们进入m.a.的土地所有者。Beaucher,肯尼迪。Gobeil, S. Jamieson, M. Hoekstra, C. McCallum, S. Roy和R. St. Clair在战场上的出色帮助和良好伙伴。
安德森,h.p., r.a.伊姆斯和o.k.斯坦塞特。1996.不连续生境走廊:对雄性根田鼠运动的影响。应用生态学杂志33: 555 - 560。
P.贝尔和R. F.诺斯。1998.(在新闻)生境走廊是否提供连通性?保护生物学121241 - 1252。
bierregard, R. O., Jr., T. E. Lovejoy, V. Kapos, A. A. dos Santos和R. W. Hutchings。1992.热带雨林碎片的生物动力学。生物科学42: 859 - 866。
Desrochers, A.和S. J. Hannon。1997.森林鸣禽在羽化后时期的穿越间隙的决定。保护生物学11: 1204 - 1210。
A. Desrochers, S. J. Hannon和K. E. Nordin。1988.北方黑顶山雀种群的冬季生存和领地获得。雀105: 727 - 736。
埃克曼,J。1984.温带柳山雀的密度季节性死亡率和种群波动(Parus montanus).动物生态学杂志53: 119 - 134。
Fahrig, L。1997.生境丧失和破碎化对种群灭绝的相对影响野生动物管理杂志61: 603 - 610。
Fahrig, L.和G. Merriam。1994.碎片化种群的保护。保护生物学850 - 59岁。:
M.吉尔平,和I.汉斯基,编辑。1991.元种群动力学:实证和理论研究。学术出版社,伦敦,英国
哈斯,c . M。1995.鸟类在农业景观中对林间走廊的传播和利用保护生物学9: 845 - 854。
汉农,s.j.和s.e.科特里尔。1998.阿尔伯塔省农业区白杨林中的巢穴捕食:来自内部的敌人。雀115第16 - 25:。
霍布斯,r . J。1992.走廊在环境保护中的作用:解决方案还是潮流?生态学和进化趋势7: 389 - 392。
赫德,c R。1996.黑头山雀的围捕声吸引不同物种(Parus atricapillus).行为生态学和社会生物学38: 287 - 292。
Ims, r。1995.与空间结构相关的运动模式。85 - 109页在L. Hansson, L. Fahrig和G. Merriam,编辑。马赛克景观与生态过程.查普曼和霍尔,伦敦,英国。
G. Inglis和A. J. Underwood。1992.对走廊生物重要性实验的一些设计提出了意见。保护生物学6: 581 - 586。
Keitt, T. H. D. L. Urban和B. T. Milne。1997.在碎片化景观中发现临界尺度。保护生态【在线】1:4。可于互联网下载,网址为:http://www.consecol.org/vol1/iss1/art4.
Kleinbaum, D. G. L. L. Kupper和K. E. Muller。1988.应用回归分析等多变量方法.第二版。美国加州贝尔蒙特,达克斯伯里出版社。
Kotliar, n.b.和j.a. Wiens。1990.多尺度的斑块和斑块结构:异质性研究的层次框架。Oikos59: 253 - 260。
朗斯,L.和A. A. Dhondt。1994.生境破碎化对凤头山雀生长发育时间的影响Parus cristatus出生的传播。宜必思136: 147 - 152。
利马,s.l.,和l.m.迪尔。1990.被掠夺风险下的行为决策:审查和招股说明书。加拿大动物学杂志68: 619 - 640。
利马,s。L。和p。a。佐尔纳。1996.走向生态景观的行为生态学。生态学和进化趋势11: 131 - 135。
Lindenmayer, d . B。1994.澳大利亚东南部木材生产森林的野生动物走廊和减缓砍伐对动物的影响:综述。野生动物研究21: 323 - 340。
林奇,j。F。和地方检察官桑德斯。1991.鸟类物种对西澳大利亚小麦带栖息地破碎化的响应:内部、边缘和走廊。pages143 - 158在检察官桑德斯和r·j·霍布斯,编辑。自然保护2:走廊的作用。萨里·比蒂,奇平诺顿,新南威尔士州,澳大利亚。
Machtans, c.s., m.a.。Villard和S. J. Hannon。1996.利用河岸缓冲带作为森林鸟类的活动走廊。保护生物学10: 1366 - 1379。
Matthysen, E., F. Adreaensen和A. A. Dhondt。1995.五子雀的扩散距离,Sitta欧洲公司,在一个高度分散的森林栖息地。Oikos72: 375 - 381。
Matthysen, E.和D. Currie。1996.栖息地碎片化降低了分散器在五倍子幼鸟中的成功Sitta欧洲公司:领土建立模式的证据。描述生态学19: 67 - 72。
K. McGarigal和W. C. McComb。1995.在俄勒冈海岸范围内景观结构和繁殖鸟类之间的关系。生态专著65: 235 - 260。
j·M·麦克纳马拉和a·i·休斯顿。1990.脂肪储备的价值以及饥饿和捕食之间的权衡。Acta Biotheoretica38: 37 - 61。
a。O。尼科尔斯和c。r。玛格尔斯。1991.为了证明走廊的生物重要性而设计的研究。49 - 61页在检察官桑德斯和r·j·霍布斯,编辑。自然保护2:走廊的作用。萨里·比蒂,奇平诺顿,新南威尔士州,澳大利亚。
Norusis, m . J。1994.SPSS高级统计SPSS,芝加哥,伊利诺伊州,美国。
Opdam, P。1991.超种群理论与栖息地破碎化:全北极鸟类繁殖研究综述。景观生态学5: 93 - 106。
Opdam, P., R. Roppen, R. Reijnen和A. Schotman。1994.鸟类保护的景观生态学方法:将元种群概念融入空间规划。宜必思137(1)补充:s139 - 146。
铁路、肯尼迪。,M. Darveau, A. Desrochers, and J. Huot.1997.北方针叶林鸟类对栖息地空白区的领地响应。秃鹰99: 976 - 980。
J. M. Reed和A. P. Dobson。1993.行为约束和保护生物学:同种吸引和招募。生态学和进化趋势8: 253 - 256。
罗宾逊,S. K., F. R.汤普森三世,T. M. Donovan, D. R. Whitehead和J. Faabord。1995.区域森林破碎化与候鸟筑巢成功。科学267: 1987 - 1990。
罗兰,J.和P. D.泰勒。1997.不同空间尺度下寄生蜂对森林结构的响应。自然(伦敦)386(6626): 710 - 713。
主编w . L。1996.个体差异导致模拟鱼群的轨迹不同。生态模型92: 65 - 77。
桑德斯,检察官,r·j·霍布斯,编辑。1991.自然保护2:走廊的作用。萨里·比蒂,奇平诺顿,新南威尔士州,澳大利亚。
桑德斯,D. A. R. J.霍布斯和C. R. Margules。1991.生态系统破碎化的生物学后果。保护生物学5: 18 32。
赛维因,k。E。m。f。威尔逊和t。l。德桑托。1996.支离破碎的南温带雨林中林下层鸟类迁徙的栖息地障碍。雀113: 944 - 949。
Simberloff博士和J. Cox。1987.保护走廊的后果和成本。保护生物学1: 63 - 71。
Simberloff, D. A. Farr, J. Cox和D. W. Mehlman。1992.运动走廊:节约还是糟糕的投资?保护生物学6: 493 - 504。
R. Slotow和E. Paxinos。1997.种内竞争影响白冠麻雀的食物返回-捕食风险权衡。秃鹰99: 642 - 650。
A. T.史密斯和M. M.皮科克。1990.同种吸引和异居群定殖率的确定。保护生物学4: 320 - 323。
泰勒,P. D. L.法里格,K.海因和G.梅里姆。1993.连通性是景观结构的重要组成部分。Oikos68: 571 - 573。
Tischendorf, L.和C. Wissel。1997.走廊作为小动物的通道:可达到的距离取决于移动模式、边界反应和走廊宽度。Oikos79: 603 - 611。
陶德和r·j·考伊。1990.测量能量货币的捕食风险:对觅食的蓝山雀进行野外实验,Parus山雀。动物行为40: 112 - 117。
韦格纳,j。F。和g。梅里亚姆。1979.鸟类和小型哺乳动物在树林和邻近农田栖息地之间的活动。应用生态学杂志16: 349 - 357。
恩斯,j . A。1994.栖息地破碎化:岛屿与鸟类保护的景观视角。宜必思137: S97-S104。
_______ .1995.景观镶嵌和生态理论。Pages1-26在L. Hansson, L. Fahrig和G. Merriam,编辑。马赛克景观与生态过程。查普曼和霍尔,伦敦,英国。
J. A. Wiens, n.c. Stenseth, B. van Horne和R. A. Ims。1993.生态机制与景观生态学。Oikos66: 369 - 380。
威尔考夫,D. S. C. H.麦克莱伦和A. P.多布森。1986.温带生境破碎化。237 - 256页在硕士Soulé,编辑。保护生物学:研究稀缺和多样性的科学。西诺尔,桑德兰,美国马萨诸塞州。
左尔纳,p。A。和s。l。利马。1997.白足鼠的景观级感知能力:感知范围和对森林栖息地的探测。Oikos80: 51-60。
记者的地址:
科琳·卡萨迪·圣克莱尔
生物科学系
阿尔伯塔大学
埃德蒙顿,阿尔伯塔,加拿大T6G 2E9,和
森林生物研究中心
Pavillon Abitibi-Price,大学拉瓦尔
圣福伊,Québec,加拿大G1K 7P4
电话:(403)492 - 9685
传真:(403)492 - 9234
cstclair@ualberta.ca
*这篇文章的版权于2000年1月1日由美国生态学会转交给恢复力联盟。
![]()