|
版权所有©1998韧性联盟*
Adina Merenlender, Claire Kremen, Marius Rakotondratsima和Andrew Weiss, 1998。监测自然资源开采对马达加斯加马索亚拉半岛狐猴的影响。保护生态(在线)2(2): 5。可从互联网上获得。URL: http://www.consecol.org/vol2/iss2/art5/
本文的一个版本,其中的文本、图、表和附录是单独的文件,可以通过以下命令找到链接.
研究 监测自然资源开采对马达加斯加马索亚拉半岛狐猴的影响艾迪娜Merenlender1,克莱尔·克雷曼2,马吕斯Rakotondratsima3.,安德鲁·韦斯4
1加州大学伯克利分校;2保护生物学中心和野生动物保护学会;3.马达加斯加野生动物保护协会;4保护生物学中心
- 摘要
- 简介
- 背景信息和研究地点
- 方法
- 人口普查的方法
- 人口统计学
- 密度、地貌和种群大小
- 结果
- 人口普查的方法
- 样本的努力
- 人口统计学
- 密度和人口规模估计
- 讨论
- 人口与资源开采的影响
- 密度和人口规模估计
- 新颖的方法
- 对本文的回应
- 致谢
- 文献引用
监测人类活动对旗舰物种的影响是保护生物多样性的重要组成部分,因为所获得的信息对制定和调整保护管理计划至关重要。在马达加斯加的马索拉半岛,我们正在监测两种最大的原猴物种,Eulemur fulvus albifrons而且Varecia variegata rubra,在受干扰和未受干扰的森林地点,以确定森林资源的开采是否对这些物种的种群生存能力有显著影响。为了测试狐猴物种对当地村民日常开采自然资源的敏感性,我们使用一种新的普查技术,在六个研究地点比较了这两种物种的人口统计和密度。其中三个研究地点离村庄较近,因此比其他地点受资源开采的影响更大。我们对600多只灵长类动物个体的数据表明,在两年的研究期间,资源提取水平对任何物种的群体规模、繁殖力或密度都没有显著影响;然而,中国的性别比例Eulemur在更受干扰的地区,雄性可能是从不太适合的栖息地迁移过来的。计算了每个地点的人口密度和整个半岛的人口规模,并用于评估该地区新公园的设计,以确保它将足够大,以支持这些受威胁的灵长类动物的存活种群。这些估计是通过从地理信息系统获得每个研究区域的表面积来计算的。对这些物种的监测继续在公园的缓冲区地区进行,在那里资源的开采仍然是允许的。
关键词:生物多样性,缓冲区,人口普查技术,保护,Eulemur fulvus albifrons,GIS,狐猴,马达加斯加,国家公园,自然资源开采,灵长类动物,保护区管理,Varecia variegata rubra。
与大陆热带森林相比,马达加斯加的森林中大型脊椎动物相对稀少。因此,2.5公斤或以上的树栖狐猴是岛上最大的脊椎动物之一,拥有相对较大的领土(Rigamonti 1993年,Sterling 1993年,Vasey 1996年)。确保它们的生存可能有利于其他活动范围更狭窄的动植物。许多狐猴以水果为食,是森林树木的重要种子传播者(White 1989, Rigamonti 1993)和一些森林冠层花朵的传粉者(Kress et al. 1994)。狐猴也可能是猛禽和陆地食肉动物的食物来源(Langrand 1990, Mittermeier et al. 1994)。由于狐猴所扮演的生态角色,狐猴分布和数量的减少可能会改变森林群落的结构、组成和功能。由于狐猴是马达加斯加的旗舰物种,它们是一个适当的分类单元,可以帮助监测马达加斯加目前保护完整、有效的生态系统和可持续管理森林资源的努力的成功(Kremen et al. 1994)。
马达加斯加狐猴目前面临三大威胁。将森林转化为农业已经导致了栖息地的极度丧失和破碎化。马达加斯加只有大约10%的原生植被保留下来(Nelson and Horning 1993, Du Puy and Moat 1996),雨林的转换估计以每年1.5%的速度继续(Green and Sussman 1990)。选择性提取木材和非木材林产品造成的低水平生境改变也可能影响狐猴种群的生存能力。为维持生计或商业用途而选择性采伐会改变森林结构和组成,潜在地影响大型狐猴在森林中移动的能力,并移除巢穴和食用树(Johns 1992, White等,1995)。最后,许多狐猴物种也被猎杀作为食物(Nicoll and Langrand 1989, Mittermeier et al.1992),如果情况严重,可能会对种群生存能力产生影响。
在本文中,我们重点研究了林产品提取对马达加斯加东北部马索拉半岛两种最大狐猴种群的影响。图1),以帮助该地区设计一个新的国家公园和森林管理区(Kremen et al. 1998;Kremen et al。在检查中).尽管这个国家公园的建立将减少狐猴栖息地破坏和狩猎对它们的威胁,但林产品提取对狐猴的影响将继续,特别是在公园周围的森林管理区内。长期来看,由于有选择地提取自然资源而导致狐猴密度的变化,可能会对可持续管理这些森林以生产生存和商业林产品的努力产生不利影响,因为狐猴是重要的传粉者和传播媒介。因此,监测狐猴对提取的反应不仅是狐猴保护的关键因素,也是森林管理计划的关键因素,为今后几年适应性地改变计划提供信息(Walters和Holling, 1990年)。
图1所示。马达加斯加马索亚拉半岛的雨林照片。
|
简单地比较不同地点狐猴的数量不太可能让我们发现林产品提取对种群生存能力的影响。例如,更受干扰的(例如,更严重的提取)森林可能为狐猴的生存提供足够的结构和食物,但不能维持正的增长率。这种区域被称为种群下沉区,当一个物种仅依靠从更适合的栖息地中发现的相邻源种群的持续迁移而居住在低质量的栖息地时,就会出现这种区域(Pulliam 1988, Dias 1996)。如果人类居民使用的森林区域毗邻一大片未受干扰的内部森林,这一被利用的区域可能是一个水池,不断吸引动物离开未受干扰的源区域。因此,这两个地区的密度可能是相似的,并没有反映出受干扰栖息地的潜在不适宜性。如果源生境继续转变为下沉生境,例如如果自然资源的开采扩大到保护区缓冲区以外,就会对种群的生存能力造成问题。
为了观察低质量栖息地对狐猴种群的影响,还必须研究其他种群测量方法。例如,狐猴fulvussp .有一个社会制度,包括女性的哲学和男性的分散(Merenlender 1993)。因此,我们可能会认为在资源有限的地区,灵长类动物群体的规模会更小,而且会偏向雌性,因为灵长类动物群体的规模通常与食物的面积和丰度有关;食物的限制可能会减少雄性的移民(Terborgh 1983, Ganzhorn 1988)。人口汇最好通过长期人口统计研究来确定,而不是通过密度(Pulliam 1988)。
通过比较在马索亚拉半岛发现的两种最大狐猴物种的人口统计参数,研究人员验证了被干扰的森林代表狐猴种群汇聚的假设,Varecia variegata rubra(图2),Eulemur fulvus albifrons(图3),分别在靠近人类住区的三个地点和较远的三个地点。我们报告了1993 -1995年期间在这些地点测量的群体规模、组成和所有社会群体的繁殖成功率,以及对密度和人口规模的估计。这一监测方案每年继续进行,以便观察人口动态的长期趋势。
图2所示。的照片Varecia variegata rubra由大卫·哈林。
|
图3所示。的照片Eulemur fulvus albifrons由大卫·哈林。
|
的亚种Eulemur fulvus在马达加斯加的森林中大量存在(Tattersall 1982),而诉诉rubra是马索阿拉半岛特有的,密度相对较低(Lindsay and Simons 1986, Mittermeier et al. 1992, White et al. 1995)。Varecia诉rubra而且e . f . albifrons由于它们的大小和饮食不同,可能对人类资源开采造成的森林组成变化作出不同的反应。Varecia诉rubra主要是果食动物,主要依赖于少数果树(Rigamonti 1993),而大肠fulvus是一个多面手,吃各种各样的水果,花,昆虫和树叶(Overdorff 1991)。这两种狐猴都吃很多人类也收割的植物。根据对狐猴饮食的研究,我们估计狐猴和人类使用的植物种类重叠的比例约为50% (Morland 1991, Rigamonti 1993;A. Merenlender, C. Kremen, M. Rakotondratsima, A. Weiss未发表的数据)和关于人类在Masoala上提取资源的信息(Raymond 1995)。这两个物种也有非常不同的生活史策略,这将影响它们如何应对栖息地的改变(Tattersall 1982)。例如,Varecia有多个后代并使用巢穴,而Eulemur通常一次生育一个后代,并携带婴儿(Morland 1992, Overdorff 1996)。因此,将这两个物种作为指标并进行比较,将有助于更好地了解林产品提取对森林生态的影响。
背景信息和研究地点马达加斯加是生物多样性保护的12个优先国家之一(Myers 1988),支持高度独特的动植物群,在许多分类组中物种级别的地方性超过75% (Nicoll和Langrand 1989年,Glaw和Vences 1994年,Mittermeier等1994年,Dransfield和Beentje 1995年)。认识到其生物多样性在国内和全球的重要性,马达加斯加政府启动了一项雄心勃勃的保护计划,提议建立14个新的保护区(世界银行1988年)。马索亚拉半岛,长4625公里2该地区约75%为原始雨林,因为它是马达加斯加现存森林面积最大的地区之一。一个2100公里的公园2于1995年向马达加斯加政府提出,并于1997年签署为法律(图4).该公园是综合保护和发展项目的一部分(Wells et al. 1992, Wells and Brandon 1993, Kremen et al. 1994);该项目与当地人民一起保护这1000公里2保护区以外的森林作为可持续天然林管理的采掘保护区。
图4所示。马达加斯加马索亚拉半岛的地图和研究区域的位置。
|
研究地点位于马索拉半岛西海岸,位于安巴尼扎纳村和坦波罗点(图4).该地区横跨13公里的海岸线,在距海岸3公里的范围内,地势崎岖,朝西,地势陡峭,从海平面上升至海拔700米(图5).我们选择了6个林区,其中一半远离村庄,因此很少或没有人参观,另一半毗邻村庄,定期接受当地人的参观,他们收获了各种各样的林产品(Raymond 1995)。除了与村庄的距离之外,小径的存在是区分受影响森林地区和未受影响森林地区的另一个标准,这表明该地区有规律的人类活动。未受干扰地点的步道系统非常有限或没有预先存在的步道系统,而被分类为受干扰地点的步道系统使用良好。我们正在量化当地村民沿着小径在距离村庄或近或远的不同距离从森林中移走的资源的数量和类型。然后,我们将能够将这些数据与定期提取的关于自然资源的村庄调查的信息进行比较,从而编制出林产品提取的完整图景。虽然每个地点用于普查的步道长度和面积不同,但未受干扰和受干扰森林地点的步道总量(16.7公里和16.8公里)和面积(1462 ha和1312 ha)几乎相同。
图5所示。每个研究地点的地图。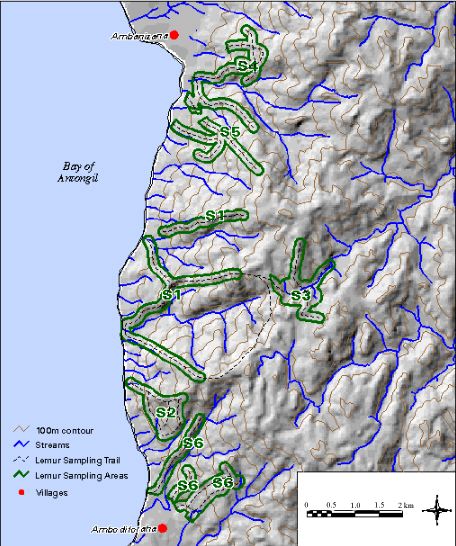
|
尽管当地村民捕猎狐猴,尤其是在农田和森林地区,但这种捕猎在我们的研究地点并不常见。因此,人类直接迁移动物并不是经常发生的事件,因此,极有可能不是灵长类动物在不同地区的人口分布或密度差异的原因。今后,研究人员的继续存在、综合保护和发展项目以及建立国家公园将有助于制止这些非法行为。
所有站点均位于低地湿润常绿森林中,以Canarium(橄榄科),Diospyros(柿科),Symphonia(金丝桃科),Tambourissa(檬立木科),Weinmannia(南蔷薇科)Anthostema而且Uapaca(大戟科)等(Rakotondratsima 1995, Kremen et al. 1998)。表1展示每个站点的物理特征。在六个研究地点中,有五个的海拔范围和地形非常相似。遗址S3是唯一位于南北陡坡以东、位于半岛内陆森林中的研究遗址。这导致该地点的平均高程升高,高程范围减小,平均向度也有所不同(表1).
表1。所有研究地点的地貌特征。
a.与最近村庄的距离。 |
我们使用的灵长类动物普查技术与经典方法(Southwick和Cadigan 1972年,Struhsaker 1975年,Defler和Pintor 1985年)在许多方面有所不同,并且允许低成本、长期的监测。我们的方法依赖于(1)反复识别社会群体,(2)获取所有已识别群体的人口统计数据。因此,该方法允许我们获得大量可靠的人口统计数据,许多地点的许多群体,因此是监测灵长类动物数量和人口统计学变化的合适方法。
1993年,选定地点,每隔50米标记小径,对两名当地村民进行了数据收集方面的培训e . f . albifrons而且诉诉rubra在每个地点进行鉴定。但是,由于1993年的数据是在培训和初步普查期间收集的,并且在该年年底又增加了两个地点,因此我们提出1993年的数据仅用于样本努力的分析。1994 - 1995年的人口和密度数据是在查明所有群体和实地工作人员得到充分培训之后提出的。
在大多数地点,采样都是在已经存在的小径上进行的,这些小径要么是村民为森林产品提取而开辟的,要么是在该地区工作的研究人员开辟的。大多数原先存在的痕迹都是沿着山脊线发现的。在普查狐猴群体时使用山脊线的踪迹是有利的,因为从山脊顶部可以很好地听到叫声,下面的斜坡的能见度通常更好。为了更好地穿过森林覆盖的山坡,有些小径确实是垂直于山脊线的(图5).在这一地区,利用已有的步道也是可取的,因为砍伐新的步道可以刺激在以前未开发的地区提取更多的林产品。
从清晨到午后,每条足迹都有两名寻找动物的人以缓慢的步伐跟随。在人口普查期间,实地工作人员试图找到所有可检测到的群体,并尽可能地计数、性别和确定年龄等级。一旦有一群组成相似的人,或有可辨认身份的人,三次出现在大致相同的地方,就会给他们一个识别号码;后来看到这群人的记录是用这个群号记录的。每次观察时收集的数据包括:(1)群体数量,(2)观察到的群体总规模,(3)成年、幼、幼雌性和雄性的数量,(4)最近的踪迹标记,(5)从踪迹标记到发现动物的地方的大致距离,(6)包括行为观察和喂养记录的注释。这些数据是1994年在每个场址为期3天的时间内从5次到8次采集的。根据1994年完成的工作,确定收集数据的当地实地技术人员对所监测的群体极为熟悉,因此可以将继续的人口普查限制在出生季节之后的几个月内,而不会失去准确性。因此,1995年访问这些场址的次数仅为2至6次,视前往场址的困难而定。
这些普查方法不同于传统的样线取样方法(Whitesides等人,1988年),因为观察者定期离开踪迹,在距离踪迹100米的范围内识别狐猴群体,以获得关于群体规模和组成的最佳数据。在田间季中期(1995年12月- 1996年1月),在两个干扰点(S4和S6)和两个未干扰点(S2和S3),将我们的方法与标准样线取样进行了比较。这样做是为了评估使用我们的现场方法计算的数据的质量和准确性,与更标准的灵长类动物普查技术进行比较。横断面法的数据是在试验中采集的(横断面),并比较两种普查技术之间的以下计算:(1)观察到的个体数量,(2)总群体大小,(3)性别比,(4)密度。
人口统计学从重复人口普查数据估计的主要人口统计参数是群体规模、性别比和女性繁殖力。确定鉴定出的群体大小和组成e . f . albifrons在每年的比较中使用了野外季节记录的最大群体规模和当时的群体组成。这样做是为了避免使用不完全的群体计数,可能导致恶劣的天气或浓密的树冠。被调查个体的性别被记录e . f . albifrons.为诉诉rubra,每个成员经常在一起的子组被确定为一个单独的组。我们确实观察到,相邻的亚群体有时会形成更大的社会群体,这一点已经在Varecia其他工人(Morland 1991, Rigamonti 1993)。我们对研究地点的大多数访问都是在旱季进行的,因为在旱季更容易观察狐猴。在这个季节,诉诉rubra通常在2 - 4只动物的小群体中观察到。我们假设这些是更大的、有凝聚力的社会群体的子集,因此,我们无法确定社会群体的规模诉诉rubra。我们也无法确定该物种的性别比例,因为区分个体的性别诉诉rubra在实地使用快速普查方法是不可靠的。
平均女性生育能力e . f . albifrons每个社会群体的出生婴儿数量除以被认为超过两岁的女性数量。因为我们无法确定成年人的性别诉诉rubra,我们假设了一个均匀的成人性别比,就像在其他研究中观察到的那样Varecia人口(河1992)。然后计算每一组的繁殖成功率,即该组出生的婴儿总数除以该组成年人数量的一半。Varecia婴儿在10月左右的旱季出生,出生后在巢中停留约3个月(Morland 1990)。Varecia诉rubra在12月以前的人口普查中一般没有观察到婴儿;因此,1993年的婴儿在1994年初首次观察到,1994年的婴儿在1995年初首次观察到。
非参数方差分析和配对检验用于检验所有研究地点的人口统计学参数。虽然嵌套anova(干扰为第1级,站点为第2级)是首选方法,但检测到显著异方差。因此,本文采用Kruskal-Wallis方差分析和Wilcoxon匹配对符号秩检验代替参数检验。这一选择并不影响研究结论。
密度、地貌和种群大小动物密度的计算方法是将每个地点所有已确定的社会群体中的成年和幼年动物的数量除以研究地点的面积。利用狐猴普查路径的GIS覆盖度和研究区域的数字高程模型(DEM)估算研究场址面积。该DEM的水平分辨率为30米,使用Arc/Info 7.1.1 TOPOGRID命令(ESRI 1996, Hutchinson 1996)开发,输入25米等高线、点高程、海岸线和数字化的马达加斯加1:10万比例尺地图水文。使用全球定位系统(GPS)绘制步道地图,并参考地形、水文、指南针和高度计读数(图4).更正全球定位系统读数需要这些额外的资料,因为马索阿拉没有基站数据,无法对全球定位系统数据进行差别更正。使用Arc/Info 7.1.1 GRID命令PATHDISTANCE (ESRI 1996)在给定的距离上缓冲这些步道,该命令计算每个位置到步道的最小距离,考虑到坡度距离。缓冲区内的表面积是通过除以每个单元的平面面积(900米)来计算的2除以斜率的余弦。
我们通过使用在类似栖息地记录的估计home range来确定用于缓冲采样轨迹的距离。Eulemur fulvus鲁弗斯在100公顷(重叠领土)的总范围内(Overdorff 1991, 1996)有40公顷(非重叠领土)的核心家园范围(Overdorff 1991, 1996)。如果核心和总住宅范围被建模为圆形区域,那么它们的半径为356 - 564米。为此,我们在350米和500米的距离上缓冲了小径E.f. albifrons,并根据这些缓冲区内的表面积估计了一个上密度和下密度。类似地,我们使用了公布的家庭范围大小的估计Varecia来自两项关于马索拉半岛的研究(Rigamonti 1993, Vasey 1996)。两项研究的住宅范围估计由24公顷至57.7公顷不等,或圆周半径为89 - 428米。因此,我们使用100米和500米的跟踪缓冲区来计算的上下限密度估计Varecia.然后利用最大缓冲距离(500 m)从地形覆盖计算研究地点的地形特征,如高程和坡向。
为了计算整个Masoala半岛和未来公园内两种狐猴种群总数的最保守估计,基于500米缓冲区的密度估计乘以在半岛或公园内发现的森林面积估计。在这些计算中使用了原始森林的整个区域,因为这两种物种已经在所有已知出现在Masoala的原始森林类型中被观察到,并且在整个半岛普遍存在(Sterling和Rakotoarison 1998年)。马索拉半岛的森林面积是根据1991年的SPOT全色图像(10米分辨率)计算出来的,该图像很容易将森林砍伐地区与原始林区区区分开。然后,根据100米等高线的数字高程模型,计算出整个半岛和核心公园区域的原始森林表面积。
我们的普查方法比标准的样条线方法具有更高的狐猴个体检出率,这是收集人口统计数据的一个优势。在密度估计上也有差异e . f . albifrons由于使用了不同的技术。利用我们的普查方法(t= 3.05, df = 18,P< 0.01) (e . f . albifrons:我们的方法N= 240,样线N= 186;诉诉rubra:我们的方法N= 84,样线N= 75)。组的规模也明显较大e . f . albifrons使用我们的方法(t= 3.98, df = 28,P<0.01)(均值= 8.28,我们的方法;均值= 6.41,样线法)。横断面法的目的是根据横断面的观察周期和长度,利用估计的普查下降距离来估计密度。采用下降距离为45.8 m的样线法,得到的种群密度e . f . albifrons106人/公里吗2;为诉诉rubra,是7.12个人/公里2,降落距离为300米。
样本的努力图6和图7显示了在1993年最初普查期间在每个地点发现的狐猴群体所需要的访问次数,在整个研究过程中多次进行普查。实地工作人员确定了77 - 100%的社会团体e . f . albifrons到第五次到访时(图6)和62 - 100%的社会群体诉诉rubra(图7).对于任何一个物种,都需要在六个地点中的一个地点进行八次访问,以确定所有的社会群体。
图6所示。样本努力曲线Eulemur fulvus albifrons,展示在连续访问研究地点后所确定的社会群体数量。
|
图7所示。样本努力曲线Varecia r . rubra,显示了在连续访问研究地点后确定的社会群体数量。
|
人口统计学
我们发现了大约428个人e . f . albifrons在1994年和1995年分别对6个研究地点的45个社会群体(表2).我们确定了48诉诉rubra1994年共有185人,1995年共有188人(表2).Kruskal-Wallis方差分析显示,干扰的总体影响不显著(1994:H(1,N= 45) = 0.56,P= 0.45;1995:H(1,N= 0.18;P= 0.67)或研究地点e . f . albifrons集团规模(1994:H(5,N= 0) = 0P= 0.75;1995:H(5,N= 0) = 0P= 0.74)。从1994年到1995年,在所有六个研究地点,Wilcoxon配对检验没有发现两个物种的总体群体大小有任何显著变化。
表2。被普查狐猴群体总数,被普查动物总数和群体大小(平均+1994年和1995年的六个地点各1 SE)。
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
下面的结果是e . f . albifrons雌雄同色的物种1994年唯一一个对青少年表现出明显女性偏见的地点(卡方= 3.5,df = 1,P(χ 2 = 5.5, df = 1,P< 0.02)为S5,自然资源开采水平较高。此外,当所有受干扰的地点集中在一起时,1994年出现了显著的雌性偏倚(卡方= 30,df = 1,P< 0.001;图8(卡方= 3.75,df=1,P< 0.05;图9).在受干扰较少的地点,与均匀的性别比没有显著差异。
图8所示。男性(左轴)和女性(右轴)的总数Eulemur f . albifrons1994年在三个受干扰较少的研究地点发现。星号表示性别比例明显偏离1:1P< 0.001。
|
图9所示。男性(左轴)和女性(右轴)的总数Eulemur f . albifrons1995年在另外三个受干扰的研究地点发现的。星号表示显著偏离1:1性别比(P < 0.05)。
|
平均雌性的繁殖力e . f . albifrons为每个研究地点计算(图10).Kruskal Wallis方差分析显示,干扰没有总体显著影响(1994:H(1,N= 45) = 0.15;P= 0.70;1995:H(1,N= 0.19;P= 0.66)或研究地点e . f . albifrons繁殖力(1994:H(5,N= 7.98;P= 0.15;1995:H(5,N= 8.72;P= 0.12)。同样的情况也发生在诉诉rubra扰动效应(1994年):H(1,N= 0.30;P= 0.58;1995:H(1,N= 49) = 0.00;P= 0.97)和生育力研究地点(1994年:H(1,N= 0.1.2;P= 0.94;1995:H(1,N= 49) = 40;P= 0.63)。然而,在这两年里,S3的社会群体的平均生育率比其他地点的低。从1994年到1995年,大多数研究地点的雌性繁殖力显著下降,如Wilcoxon配对符号秩检验的Wilcoxon秩和(图10).
图10所示。雌性繁殖力值Eulemur f . albifrons在每个研究地点。生育能力是用1994年和1995年出生的婴儿数量除以被认为超过两岁的女性数量来衡量的。星号表示重要结果(P年份间Wilcoxon配对的<0.07)。
|
生殖率为诉诉rubra1994年不受研究地点的影响(df = 5,N= 49,H= 1.22,P= 0.94)或1995 (df = 5,N= 48岁H= 3.43,P= 0.63)。在1993年至1994年期间,繁殖能力没有显著增加或减少的趋势诉诉rubra在所有研究地点(图11).然而,一个种群很少连续数年繁殖。在这两年在所有六个地点进行普查的48个群体中,我们记录了17个在1993年只有婴儿的群体,20个在1994年只有婴儿的群体,4个在两年都有婴儿的群体,7个在任何一年都没有婴儿的群体。
图11所示。雌性繁殖力值Varecia诉rubra在每个研究地点。生育能力是用1994年和1995年出生的婴儿数量除以被认为超过两岁的女性数量来衡量的。
|
密度和人口规模估计
的低和高平均密度e . f . albifrons对于六个站点,基于两个跟踪缓冲区(参见方法),范围为15.67至21.52只/公里21994年为16.56 ~ 22.73头/公里2在1995年(表3).为诉诉rubra,密度从每公里7.36到27.28只不等21994年为7.56 ~ 28.02只/公里2在1995年。
表3。最小和最大密度(不。动物/公里2)e . f . albifrons而且诉诉rubra在1994年和1995年在马索亚拉半岛的六个研究地点发现的,这是根据采样轨迹周围的可变缓冲区宽度计算出的面积估计的。
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
干扰点与未干扰点的密度差异不显著e . f . albifrons或诉诉rubra1994年或1995年期间的最低或最高估计数。配对t1994年至1995年的测试显示,各年之间的密度估计有显著差异(e . f . albifrons350年的今天,米缓冲区:t年代= 2.9, df = 5,P< 0.05;e . f . albifrons500年的今天,米缓冲区:t年代= 2.9, df = 5,P< 0.05;诉诉rubra,100年的今天,米缓冲区:t年代= 3.7, df = 5,P< 0.05;诉诉rubra,500年的今天,米缓冲区:t年代= 4.29, df = 5,P< 0.05)。为e . f . albifrons从1994年到1995年,所有地点的密度都有所增加诉诉rubra并没有在所有网站上显示出相同的趋势。
1995年最保守的平均值(+1 SE)密度估计,基于500 m缓冲区,结果估计的人口规模诉诉rubra16179年+3254年和35451年+4715年e . f . albifrons仅在公园内,就有25542人+5137年和55967年+整个森林面积分别为7444。
Eulemur是马达加斯加分布最广的原猴属之一;其成员被认为能够适应中等程度的森林干扰(Tattersall 1982)。我们的结果为e . f . albifrons支持这一观点,因为我们只能在邻近村庄的森林地区和马索亚拉西海岸未受干扰的地点之间检测到群体规模和组成的微小差异。相比之下,其他人则是这么认为的Varecia对森林健康敏感(White等,1995年),因此可能受到自然资源开采的影响。这一概念在我们的研究中没有得到支持诉诉rubra,因为我们无法从现有数据中区分受干扰和未受干扰森林地区的雌性繁殖力和密度,尽管我们的监测项目中没有包括显著变化的森林。马达加斯加的一些森林已经受到了影响,狐猴几乎没有生存的空间。可能需要更多年份的数据,以查明受保护森林斑块与利用森林斑块之间繁殖成功的显著趋势。然而,我们的数据表明,像马索拉这样低水平的森林资源开采可能对狐猴种群没有什么威胁。
这些初步结果是令人鼓舞的,因为只要保持森林覆盖和禁止捕猎狐猴的禁令,公园外留下的森林区域(123,865公顷)很可能继续为狐猴种群提供栖息地。这些区域被故意留在公园外,作为公园的缓冲区;在这一地区,木材和非木材森林产品的销售作为农业扩张的一个有利选择正在被推广(Kremen等人,在检查中).如果不采取措施增加现存森林对当地人的价值,并在经济上为他们提供支持,农业扩张最终不仅会破坏这些边界森林,还会破坏公园内的森林,就像在马达加斯加其他地方发生的那样(Smith et al.1997)。利用这些边境森林进行强化开采可能最终会降低它们对狐猴和其他生物的栖息地适宜性,但它为整个半岛的整体保护和资源管理战略提供了一个关键因素。在这些和其他研究地点对这两种狐猴的监测仍在继续,以评估保护计划的有效性和边境森林中加强采伐的影响。
人口与资源开采的影响为了评估保护区在保护灵长类动物方面的价值,以及监测生境管理活动对灵长类动物的影响,包括在不受保护地区或开采保护区的资源开采,需要跨时空的灵长类物种可靠的人口统计数据。灵长类动物的人口统计变量通常在不同地点、栖息地类型、社会群体和个体之间具有高度的变化(Altmann et al. 1985, Richard et al. 1991, Sussman 1991);然而,许多灵长类动物的研究只专注于一个或少数群体,因此在文献中很少有统计上可靠的人口参数估计(Dobson and Lyles 1989)。这是不幸的,因为人口统计数据更有可能揭示栖息地管理活动对灵长类种群的影响。
灵长类动物群体的大小通常与资源分布和丰度有关(Terborgh 1983, Ganzhorn 1988, Chapman 1990, Janson 1992)。当可用资源有限时,在受干扰森林或边缘森林中发现的灵长类动物群体可能更小,而当再生森林产生更多的食物资源时,灵长类动物群体可能更大(Ganzhorn 1988, Olupot et al. 1994)。然而,对我们初步结果的分析并没有发现受村民自然资源开采影响的研究地点之间的群体规模有显著差异。
雌性繁殖能力因个体、社会群体、年龄和栖息地而异。关于大范围内许多个体狐猴物种的繁殖率的信息很少(Sussman 1991)。因此,我们的研究代表了野外狐猴繁殖力最全面的监测工作之一。在这项研究中,我们没有发现人类干扰对两个物种的繁殖力的影响。这表明,个体和社会群体的差异解释了更多的生育率的变化,而不是栖息地完整性的差异或森林类型的差异,这是本研究的重点。在e . f . albifrons我们发现,大多数地点的雌性繁殖力存在显著的年际差异。这支持了早期的一项研究,该研究表明,不同年龄的女性个体的出生率存在差异Eulemur在马达加斯加的其他地方(Overdorff等,在检查中).同样,我们的研究结果也支持了先前的一个建议Varecia在野外每两年只成功繁殖一次(Morland 1990)。这一出生间隔可能是由于Varecia经常一次生育几个婴儿,并经常成功地在野外抚养一个以上的婴儿,这需要比那些在生育间隔为一年的情况下抚养一个婴儿的原猴动物消耗更多的能量(Tattersall 1982)。
可用的食物资源和森林结构可以影响人口统计参数,如存活率和扩散模式,雄性和雌性之间的差异,从而改变灵长类种群的性别比例。虽然我们没有足够的数据来统计分析婴儿性别比例,但我们从圈养研究和其他实地研究中知道大肠fulvus(Watson et al. 1996, Overdorff et al. 1991, Overdorff et al.,在检查中).性别间死亡率和人口分布的差异影响着老年阶层的性别比例。因此,我们认为,在受干扰地点的老年阶层中,女性偏倚的性别比例可能是由于相对于男性移民来说,更多的男性居民分散,或者是相对于女性的男性死亡率增加。在Eulemur,雌性通常是多情的,而雄性则分散;因此,我们假设,在资源不足的地区,雄性会从资源不足的地区分散,导致观察到的雌性偏倚的性别比例。相比之下,没有证据支持在受干扰地区男性和女性之间的死亡率差异。
在这些早期的监测工作中,除了在受干扰的地点发现雌性狐猴性别比例偏向的趋势外,我们无法发现邻近农村的森林地区和那些较远的地区狐猴数量的显著差异。我们犯第二类错误的概率并不高:基于我们1994年数据的力量估计在群体规模和繁殖力数据上都很高。对两种狐猴来说,检测群体规模增加或减少4%或以上的能力都大于0.97,而对两种狐猴来说,检测雌性生殖力变化4%或以上的能力都大于0.9。使用monitor.exe计算的I型错误率为0.05 (Gibbs 1998)。灵敏度分析表明,这种功率分析的结果对我们提供的一些估计输入值不敏感(例如,每次站点访问观察灵长类群体的次数)。其他研究通常接受的功率水平只允许检测到20%的增加或减少的高概率(Zielinski和Stauffer, 1996)。即使是被深入研究的加州斑点猫头鹰(思occidentalis)在检测到4%的人口规模变化时,只产生了0.13到0.64之间的功率估计(Taylor and Gerrodette 1993)。
密度和人口规模估计我们没有发现人力资源利用对狐猴密度的显著影响。然而,这些结果是基于很短的时间内,受干扰较多的森林遗址也毗邻大量未受干扰的森林。我们怀疑该地区为这两种狐猴提供了重要的种群来源,因此很难检测受干扰程度较高和较低的地点之间的总体密度差异(Dias 1996)。
我们计算的人口密度估计e . f . albifrons与我们的方法相比,用我们和Sterling和Rakotoarison(1998)测量的马索阿拉半岛上的条形线普查方法发现的结果要低,或为大肠fulvussp.马达加斯加其他地方(Morland 1991, Sterling and Rakotoarison 1998, Overdorff et al.,在检查中).为诉诉rubra,最高平均密度估计(基于100米缓冲区)与Sterling和Rakotoarison(1998)在马索拉东部发现的几乎相同。
我们对整个半岛狐猴物种和国家公园狐猴物种数量的估计是基于这样一个假设:半岛上的所有森林地区都为这些物种提供了合适的栖息地。在整个半岛的田野工作者都能看到这两种物种;然而,考虑到由于栖息地类型的不同而导致的动物数量的不同水平,这些估计可以通过来自半岛其他地方的数据得到改进。
我们得出的人口规模估计与直接从公布的家栖范围内推断出的总人口规模在同一个数量级上。使用100公顷和40公顷的上、下住宅面积e . f . albifrons(Overdorff 1991),根据我们1995年的数据,每组平均规模为9.39只动物,e . f . albifrons如果所有森林地区都被社会群体完全占领,马索拉半岛的动物数量将达到31,740 - 79,345只。我们的人口规模估计(平均+1 SE),整个半岛为55967+7444只动物。类似地,使用上下限家庭范围估计与对应的平均群体规模诉诉rubra根据已发表的研究(Rigamonti 1993, Vasey 1996),我们估计半岛上的动物数量为75,725 - 144,020只。我们估计整个半岛的人口规模为25,542人+5137只动物,更保守的估计。我们利用这些种群数量估算来评估拟议中的马索拉国家公园的设计,得出结论认为,拟议中的公园有214,145公顷的森林面积,足以保护每个物种的大量种群。
新颖的方法鉴于世界上超过90%的灵长类动物濒临灭绝,开发有效的监测灵长类动物的方法是灵长类动物保护生物学的重要一步。为了监测狐猴种群,我们采用了几种新的田野和分析方法。研究人员开发了一种低技术含量的狐猴社会群体识别方法,用于长期监测,以最大限度地收集与灵长类动物人口学相关的数据。然而,由于没有标记单个动物,如果观察人员集中或分割群体,可能会引入一定程度的错误。为了测量这种误差以保证质量,我们必须标记几个灵长类动物群体,以便将标记动物和未标记动物的数据进行比较。这需要大量的动物处理,这是本项目无法完成的一部分。然而,综合保护和开发项目将大大受益于有效的质量控制方案。这样的程序将提供对生物和社会经济监测的误差水平的估计,这些误差是由项目中不同参与者进行的数据收集中固有的。例如,美国的一个森林健康监测项目测量了有经验的训练人员和收集物种丰富度和覆盖数据的无经验技术人员之间的一致性,发现技术人员的数据非常精确(Stapanian等,1997年)。
一个由当地村民组成的团队在我们的培训和监督下收集了所有的实地数据。这些方法充分利用了土著人民对森林动植物的知识。在美国,人们普遍认为环境质量监测应该由当地公民参与,这导致了志愿者监测项目的增加(Kerr et al. 1994)。通常,监测大量自然资源(如区域水质)的工作需要大量的数据收集者,而这些数据只能通过志愿者计划获得(Heiskary et al. 1994)。正如我们的研究所证明的那样,经过培训的当地居民不仅可以提供有价值的数据,而且收集生态数据的行为也增加了他们对周围生态系统的理解,以及可持续土地利用实践的重要性。这是非常宝贵的,因为我们的环境的健康不应只取决于科学家和自然资源管理者。
我们的方法比传统的灵长类动物行为观察技术需要更少的现场时间,但比快速调查方法更密集。传统的野外灵长类动物学强调动物个体行为和社会群体动态的重要性,这需要研究已知的(通常被标记的)个体,因此极大地限制了动物和研究地点的数量(Dobson和Lyles 1989)。灵长类动物密度的快速调查(Southwick and Cadigan 1972, Struhsaker 1975, Defler and Pintor 1985, Buckland et al. 1993, Sterling and Rakotoarison,在新闻)也极具局限性,因为灵长类动物的密度通常很低,而且往往难以捉摸,因此限制了短期实地访问中可能观察到的数量。我们描述的人口普查方法最大限度地提高了以下能力:(1)在大量研究地点研究灵长类动物的许多社会群体,(2)反复识别狐猴居住群体,以及(3)量化社会群体组成和密度。与传统的样线法相比,这些新方法也提高了检出率。使用我们的普查方法,我们观察了更多的个体,记录了更大的群体规模,从而获得了更多的人口统计数据。尽管检出率较高,但我们的密度估计低于其他研究人员在Madagscar发现的结果(Morland 1991, Sterling和Rakotoarison 1998, Overdorff et al.)。在检查中).这表明横断面估计是不切实际的高,特别是对e . f . albifrons。密度的估计诉诉rubra,使用我们的方法,与使用样条线方法和其他研究者(Sterling和Rakotoarison 1998)计算的结果相似。这些差异表明估算森林灵长类动物种群密度的固有困难。
这里描述的重复人口普查技术对于在空间和时间上监测灵长类动物种群非常有用,可以提供种群参数的统计估计和估计种群规模。地理信息系统的使用可以对研究地点进行扰动、地貌和面积方面的表征,也可以摆脱测量的样线取样的限制,这种限制严重限制了对动物(如狐猴)的实地观察,因为狐猴在大片地区呈斑块状分布。利用地理信息系统还可以将人口规模的估计外推到更大的地区。这些方法可以用于监测其他有领地的动物。这些方法的结合使我们能够在支持资金有限的情况下,在几个研究地点估计两种狐猴的多个社会群体的种群密度,并跟踪它们的人口趋势。对这些参数的可靠估计可以用来比较备选的生境管理方案,从而为适应性管理过程提供输入(Walters和Holling 1990年,Kremen等1994年)。
欢迎对本文进行回复。如果被接受发表,您的回复将被超链接到文章。要提交评论,请关注这个链接.要阅读已接受的评论,请关注这个链接.
致谢
奥特曼,J., G.豪斯法特,S. A.奥特曼。1985.安博塞利狒狒的人口统计学。美国灵长类动物学杂志8: 113 - 125。
查普曼,c。1990.三种新热带灵长类动物群体大小的生态学相关性。叶形线Primatologica55: 1 - 9
普里勒,t。R。和d。品托。1985.在已知灵长类动物密度的森林中,用样带对灵长类动物进行普查。国际灵长类动物学杂志6: 243 - 259。
迪亚斯,p . C。1996.种群生物学的源和汇。生态学与进化生物学趋势“,11(8): 326 - 329。
多布森,a.p.,和a.m.莱尔斯。1989.灵长类动物的种群动态和保护。保护生物学3.: 362 - 380。
德兰斯菲尔德,J.和H.本杰。1995.马达加斯加的棕榈树。英国伦敦皇家植物园,邱园和国际棕榈协会。
杜佩,D. J.和J. Moat。1996.基于基础地质对马达加斯加原始植被的精细分类:利用GIS绘制其分布并评估其保护状况。205 - 218页在w·r·洛伦科,编辑。Biogeographie de马达加斯加。ORSTOM,巴黎,法国。
环境系统研究所。1996.Arc/Info版本7.1.1在线文档。雷德兰兹,加州,美国。
Ganzhorn j . U。1988.马达加斯加灵长类动物之间的食物分配。环境科学75: 436 - 450。
吉布斯,J。1998.监控程序:Monitor.exe
http://www.im.nbs.gov/powcase/powcase.html
格劳和万斯先生。1994.马达加斯加两栖动物和爬行动物的野外指南。第二版。私人出版,德国科隆。
格林,g.m.和R. W.萨斯曼。1990.从卫星图像看马达加斯加东部雨林的砍伐历史。科学248: 212 - 215。
海斯卡里,S.林德布鲁姆和C. B.威尔逊。1994.利用公民志愿者数据检测水质趋势。湖泊和水库管理9(1): 4 - 9日。
哈钦森,m F。1996.数字高程模型插值的局部自适应方法。第三届综合地理信息系统和环境模型国际会议/讲习班论文集新墨西哥州圣达菲,1996年1月21日至26日。圣巴巴拉,加利福尼亚州,美国。
詹森,c . H。1992.灵长类动物社会结构的进化生态学。95 - 130页在e·a·史密斯和b·温特海尔德,编辑。进化生态学和人类行为。Aldine de Gruyter,纽约,纽约,美国。
约翰,a D。1992.热带雨林的物种保护。页面15-53在t·c·惠特莫和j·a·塞耶,编辑。热带森林砍伐和物种灭绝。查普曼和霍尔,伦敦,英国。
科尔,M., E. Ely, Y. Lee和A. Mayio。1994.环境监测志愿者简介:全国调查结果。湖泊和水库管理9(1): 1 - 4。
克莱门,C., A. M.梅伦德,D. D.墨菲。1994.生态监测:热带地区综合保护和发展项目的重要需要。保护生物学8: 388 - 397。
克莱门,c·K, i·雷蒙德,K·兰斯。1998.监测马达加斯加保护影响的跨学科工具。保护生物学12: 549 - 563。
克莱门,C., V. Razafimahatratra, R. P. Guillery, J. Rakotomalala, A. Weiss,和J. Ratsitsompatrarivo。在审查。在科学上:设计马达加斯加的马索拉国家公园。
Kress, W. J. G. E. Schatz, M. Andrianifahanana和H. S. Morland。1994.授粉的Ravenala Madagascariensis(Strelitziaceae)发现的马达加斯加狐猴:古老共同进化系统的证据。美国植物学杂志81: 542 - 551。
Langrand, O。1990.马达加斯加鸟类指南。耶鲁大学出版社,纽黑文,康涅狄格州,美国。
林赛,n.b.d.,和h·j·西蒙斯。1986.笔记Varecia在其活动范围的北部界限。渡渡鸟23: 19到24。
Merenlender, a . M。1993.社会性对人口和遗传结构的影响狐猴fulvus鲁弗斯(一夫多妻)和狐猴rubriventer(一夫一妻制)和保护意义。论文。美国纽约州罗切斯特罗切斯特大学。
米特迈耶,R. A.康斯坦,M. E.尼科尔和O.兰格兰。1992.马达加斯加狐猴:1993-1999年狐猴保护行动计划。世界自然保护联盟、腺、瑞士。
密特迈耶,R. A.塔特萨尔,W. R.康斯坦特,D. M.迈耶斯,R. B.玛斯特。1994.马达加斯加的狐猴。保护国际,华盛顿特区,美国。
河,h·S。1990.皱狐猴亲代行为与幼猴发育(Varecia variegata)在马达加斯加东北部的雨林中。美国灵长类动物学杂志20.: 253 - 265。
_________。1991.狐猴社会组织初步报告(Varecia variegata variegata)在马达加斯加东北部的雨林中。叶形线Primatologica56: 157 - 161。
_________。1992.马达加斯加诺西曼加布低地雨林黑白和白条纹狐猴的社会组织和生态。论文。耶鲁大学,纽黑文,康涅狄格州,美国。
迈尔斯,N。1988.受威胁的生物:热带森林中的“热点”的环保主义者8: 1。
纳尔逊,R.和N.霍宁。1993.AVHRR-LAC对马达加斯加森林面积的估计,1990年。国际遥感杂志14: 1463 - 1475。
尼可尔,m.e.和欧·兰格朗。1989.马达加斯加:自然保护讽刺剧protégées。世界自然基金会,瑞士格兰德。
Olupot, W. A. Chapman, C. H. Brown和P. M. Waser。1994.白眉猴(Cercocebus albigena)人口密度、群体规模和范围:20年的比较。美国灵长类动物学杂志32: 197 - 205。
Overdorff d J。1991.生态与两种原猿灵长类动物的社会结构相关:Eulemur fulvus鲁弗斯而且Eulemur rubriventer在马达加斯加。论文,杜克大学,达勒姆,北卡罗来纳州,美国。
_________。1996.马达加斯加两种狐猴的生态与社会结构相关。美国体质人类学杂志One hundred.: 487 - 506。
Overdorff, d.j., A. Merelender, P. Talata, A. Telo, Z. Forward。在审查。人口统计学和生活史Eulemur fulvus鲁弗斯从1988年到1995年在马达加斯加东南部。
·普廉姆h·R。1988.源、汇和人口调节。美国博物学家132: 652 - 661。
雷蒙德,我。1995.植物生态学的方法,评价定性和定量利用的村庄,自然资源forêt密集潮湿的永恒。DEA论文。塔那那利佛大学,塔那那利佛,马达加斯加。
理查德,A. F., P. Rakotomanga和M. Schwartz。1991.人口统计学的Propithecus verreauxi在马达加斯加的Beza Mahafaly: 1984-1988年的性别比例、存活率和生育率。美国体质人类学杂志84: 307 - 322。
Rigamonti, M . M。1993.红冠狐猴的活动范围和饮食(Varecia variegata rubra)在马达加斯加的马索拉半岛。25 - 39页在p.m. Kappeler和j.u. Ganzhorn,编辑。狐猴的社会系统及其生态基础。全会出版社,纽约,纽约,美国。
史密斯,A. P. N.霍宁,D.摩尔。1997.马达加斯加西部区域生物多样性规划和狐猴保护与GIS。保护生物学11: 498 - 512。
索斯威克,c。H。和f。c。卡迪根。1972.马来西亚灵长类动物的种群研究。灵长类动物13:队。
斯塔帕尼亚,m.a., S. P.克莱恩,D. L.卡塞尔。1997.大规模生态调查中森林植被测量方法的评价。环境监察及评估45(3): 237 - 257。
英镑、e·J。1993.咿呀语的范围使用模式与社会组织(Daubentonia madagascariensis)。页1 - 9在p.m. Kappeler和j.u. Ganzhorn,编辑。狐猴的社会系统及其生态基础。全会出版社,纽约,纽约,美国。
斯特林,E. J.和N.拉科托阿里森。1998.马达加斯加东部马索拉半岛灵长类物种丰富度和密度的快速评估。叶形线Primatologica69(suppl.1)。
Struhsaker, T . T。1975.红疣猴。芝加哥大学出版社,美国伊利诺斯州芝加哥。
苏斯曼,R W。1991.自由放养的人口和社会组织狐猴雌猫在马达加斯加的贝扎马哈法利自然保护区美国体质人类学杂志84(1): 43-58。
塔特萨尔,我。1982.马达加斯加的灵长类动物。哥伦比亚大学出版社,美国纽约。
泰勒,b·L,和t·杰罗黛特。1993.统计力量在保护生物学中的应用:小头鼠鸵和北方斑点猫头鹰。保护生物学7(3): 489 - 500。
Terborgh, J。1983.五种新的世界灵长类动物。普林斯顿大学出版社,美国新泽西州普林斯顿。
维西,N。1996.红缘狐猴的取食和放养行为(Varecia variegata rubra)及白额狐猴(狐猴fulvus albifrons).美国体质人类学杂志增刊。22: 234 - 235。
沃尔特,c。J。和c。s。霍林。1990.大规模的管理实验和边做边学。生态71(6): 2060 - 2068。
华生,S . J.沃德,K.伊泽德,D.斯塔福德。1996.圈养原猴种出生性别比偏倚分析。美国灵长类动物学杂志38(4): 303 - 314。
威尔斯,M.和K. E.布兰登。1993.缓冲区的原则和实践以及当地参与生物多样性保护。中记录22: 157 - 162。
威尔斯,M, k,布兰登,l,汉娜。1992.人与公园:将保护区管理与当地社区联系起来。世界银行出版物,美国华盛顿特区。
白色,f·J。1989.黑白皱狐猴的饮食、活动行为和社会组织Varecia variegata variegata东南部马达加斯加。美国体质人类学杂志78(2): 323。
怀特,F. J.奥弗多夫,E. A.巴尔科,P. C.赖特。1995.皱狐猴的分布(Varecia variegata)在马达加斯加的拉诺玛法纳国家公园。叶形线Primatologica64(3): 124 - 131。
怀特塞德斯,g.e., J. F.奥茨,S. M.格林,R. P.克鲁伯丹兹。1988.从西非雨林横断面估算灵长类动物密度:技术的比较。动物生态学杂志57(2): 345 - 368。
世界银行、美国国际开发署、瑞士合作组织、联合国教科文组织、联合国开发计划署和世界自然基金会。1988.马达加斯加环境行动计划。卷1。世界银行,华盛顿特区,美国。
齐林斯基,W. J.和H. B.斯托弗。1996.监控集市在加州人口;勘测设计与功率分析。生态应用程序6(4): 1254 - 1267。
记者的地址:
艾迪娜Merenlender
环境科学、政策和管理部
加州大学伯克利分校
加州伯克利,美国94720-3110
电话:(707)744 - 1270
传真:(707)744 - 1040
adina@nature.berkeley.edu
*这篇文章的版权于2000年1月1日从美国生态协会转让给韧性联盟。
![]()