

去pdf本文的版本
马托内,R. G., A. Bodini和F. Micheli. 2017。利用定性循环分析确定自然扰动和管理决策对沿海渔业社会生态系统的潜在后果。生态和社会22(1): 34。
https://doi.org/10.5751/ES-08825-220134
利用定性循环分析确定自然扰动和管理决策对沿海渔业社会生态系统的潜在后果
摘要
管理可持续发展和资源提取需要理解生态系统和人类之间的反馈。这些反馈是复杂社会生态系统(SES)的一部分,在这个系统中,资源、参与者和治理系统相互作用,在这些组成部分之间产生结果。定性建模方法为评估复杂的SES动力学提供了方法。特别是在对函数关系和参数值知之甚少的数据贫乏系统中,循环分析对于检查和识别外部扰动和管理干预的潜在结果非常有用。通过一个多物种、多船队沿海小规模渔业的案例研究,我们展示了循环分析的应用,以提供关于SES对扰动和管理行动的响应的预测。具体而言,我们研究了不同治理干预措施(例如,领土用户权利、渔场关闭、市场激励、生态旅游补贴)和环境变化对沿海渔业的潜在生态和社会经济后果。我们的结果表明,生物物理和社会经济成分之间的复杂反馈可能导致反直觉和意想不到的结果。例如,通过生态旅游或补贴创造新的就业机会可能对渔业合作社成员与非成员产生混合影响,突出了公平问题。假设生态标签对市场价格有直接影响,并且在大多数模型结构下缺乏负面的生物学影响,那么生态标签等基于市场的干预措施预计将产生总体积极的经济效应。我们的研究结果强调,在一个独特的管理单元中整合生态和社会变量,可以揭示理想的生态和社会结果之间的重要潜在权衡,突出哪些用户群体可能更容易受到外部冲击的影响,并确定应该进一步测试哪些干预措施,以确定可持续发展范式三重底线下的潜在双赢结果。介绍
越来越多的证据表明生态系统退化及其对人类福祉的相互影响,因此要求在全世界范围内进行基于生态系统的全面管理。自然资源管理已经从关注单一物种或部门、将环境视为静态的、将社会和生态问题分离的方法转向考虑整个生态系统(包括人类、社会和生态成分之间的相互作用以及多种活动的累积影响)的综合的、动态的方法(Hughes et al. 2005, McLeod et al. 2005, Leslie and McLeod 2007, Levin et al. 2009)。这种转变反映了这样一种观点,即管理可持续发展和资源提取需要理解生物物理系统和人类之间的反馈,因此需要一种综合的、跨学科的方法。这些反馈是社会生态系统(SES)的一部分,其中资源、行为者和治理系统相互作用,在这些组成部分之间产生结果(Berkes和Folke 1998, Ostrom 2009, Cox等人2010,McGinnis和Ostrom 2014)。
海洋生态系统及其所支持的渔业是复杂的SES的例子,生态、社会、经济和制度组成部分之间在多个尺度上运作着许多关系(Berkes 2006, Mahon等人2008,Levin等人2009,Berkes 2011, Halpern等人2012,Kittinger等人2013)。治理和金融系统与社会系统相互作用,影响人类行为,进而影响海洋和沿海环境,而海洋环境和资源单位又反过来影响操作和集体选择规则的选择,以及经济和文化价值(图1)。所有这些元素相互作用,产生一系列动态结果(Berkes and Folke 1998, Mahon et al. 2008, Ostrom 2009, McGinnis and Ostrom 2014)。目前的海洋资源管理认识到考虑渔业组成部分之间的相互作用以满足社会、生态和经济可持续性的重要性,但要明确理解渔业对相互作用的物种的更大网络的直接和间接影响以及这些组成部分对更大的社会-生态网络的反馈,可能是一项挑战。需要量化和建模的过程是大量的,并且产生的反馈的效果很难预测。这些复杂的动态往往超出了我们目前的理解和数据可用性。然而,为了为决策提供信息并指导未来的监测和管理行动,管理人员必须评估管理行动或系统受到的外部干扰(例如,与气候变化、市场波动有关的驱动因素)对整个系统的影响,以避免意想不到的后果并进行适应性管理。beplay竞技
一套建模框架已经开发出来,以检查耦合SES中生物和人类对多个外部和内部驱动因素的反应(MIMES, Boumans和Costanza 2007;投资,纳尔逊等。2009;亚特兰蒂斯,富尔顿等人。2011)。大多数这些模型需要大量的数据,因为系统组件数量众多,反馈也很复杂。对社会经济系统及其动态进行深入定量描述之外的另一种方法是定性建模。定性建模方法,包括模糊认知映射(Kok 2009)、因果循环图(Lane 2008)、贝叶斯信念网络(Woolridge和Done 2004)和循环分析(Puccia和Levins 1986),可以为评估外部扰动和管理策略提供实用工具。特别是在数据可用性有限、社会经济和生物物理成分之间的关系的性质和强度存在不确定性的情况下(Fishwick和Luker 1991年,Dambacher等人2007年,Espinoza-Tenorio等人2013年,Carey等人2014年)。此外,它们可以使用与多个利益相关者的参与式方法进行开发,具有概念性吸引力,直观,并代表良好的沟通工具。回路分析是一种特殊的技术,当相互作用的符号是已知的,但联系的其他方面是不确定的,包括相互作用的强度及其函数形式和参数时,它允许研究复杂系统的动力学;换句话说,一个系统组件(例如,物种、参与者或用户组)对与它交互的另一个组件有积极的、消极的或没有影响。 Despite its limitations (see discussion of merits and limitations in Justus 2006), the limited data requirements of loop analysis make it a promising tool for investigating the dynamics of SES in data poor systems.
循环分析允许检查外部压力扰动(Bender et al. 1984)如何通过变量之间的相互作用网络在系统中跨越多个社会经济和生态变量潜在地传播其影响。在循环分析中,变量之间的联系表示一个变量对另一个变量的变化率的定向影响。例如,在捕食者-被捕食者关系中,捕食者数量的增加导致其猎物的生长速度下降。这些连杆是用微分方程组的雅可比矩阵的系数在数学上表示的。这些系数的符号表明任何一个变量如何在质量上影响其他变量。通过遵循链接的方向,人们可以重建相互作用的途径,通过这些途径,管理行动或自然扰动传播到目标变量之外。在自然扰动或管理干预之后,可以预测系统中任何变量的丰度或水平将增加、减少或保持不变。
因此,循环分析为处理数据匮乏系统中SES的复杂性提供了定量建模的替代方案,并就SES对扰动和/或管理行为的响应提供了可测试的假设(例如,Carey et al. 2014)。尽管循环分析为研究海洋社会-生态系统的复杂性提供了一个分析框架(Espinoza-Tenorio等人,2013年,Carey等人,2014年,Reum等人,2015年),但迄今为止,该方法的许多应用都集中于系统的生态元素,而没有包括社会系统的各个方面和关键的SES联系(但见Dambacher等人,2007年)。
我们应用定性循环模型来研究墨西哥下加利福尼亚州维兹卡诺半岛沿岸SES的变化,这些变化是对自然扰动和最近实施或正在考虑的管理措施的响应。这些沿海渔业作为一个模范系统特别重要,因为从20世纪30年代开始,该地区的渔业合作社被授予了对一系列无脊椎动物物种的独家使用权(McCay等人,2014年),而捕捞鳍鱼或不属于合作社的渔民仍然是开放的。因此,该系统允许审查为不同物种和不同用户分配使用权的生态和社会经济后果(例如,Pomeroy等人,2001年,Costello等人,2008年),这是一种目前正在全球小规模渔业中实施的管理方法(http://www.bloomberg.org/program/environment/vibrant-oceans/).
通过对多物种、多船队、沿海小规模渔业的案例研究,我们展示了循环分析的应用,以检查与领土用户权渔业(TURF)管理的商业渔业相关的生态和社会经济变量之间的相互作用,并了解驱动这些耦合系统的社会和生态性能的主要反馈。我们演示了如何将循环分析作为一种方法,用于检查场景分析的潜在结果,从而为SES的规划和监控提供信息。通过这种方法,我们研究了一组模拟对SES扰动的情景,包括最近与观察到的不同种群下降相关的气候扰动的影响(例如,Micheli等人,2012年),以及生态标签等市场举措,生态标签被授予该地区的一个渔业(刺龙虾捕渔渔业)(Micheli等人,2014年)一个),以及目前正在考虑的渔业管理和治理的变化(Micheli等人,2014年b).虽然这些设想是为了捕捉特定于维兹卡诺渔业的扰动和管理行动,但它们代表了世界上许多沿海渔业社区所面临的影响以及其中一些社区可采用的管理方案。由于循环分析在渔业管理领域的应用相对较新(但参见Espinoza-Tenorio等人,2013年,Carey等人,2014年),我们通过对案例研究系统中的现实场景的分析,强调了循环分析在评估复杂社会生态系统方面的潜力。除了维兹卡诺地区的合作社外,我们的分析应该为一系列小规模渔业提供有用的见解,以确定监测的假设和关键变量。具体来说,我们问:(1)外部干扰导致特定种群下降的预期生物和社会经济后果是什么?(2)什么样的市场化、治理或地方管理行动可以带来生物和社会经济效益?哪些行动可能导致资源减少或负面的社会经济影响?(3)系统的表示(即系统组件之间包含哪些具体的联系和反馈)如何影响对摄动的预测响应?
方法
研究系统/概念模型
为了说明和描述Vizcaino渔业系统的组成、联系、相互作用和潜在结果,我们基于Ostrom的社会生态系统(SES)框架(Ostrom 2009, McGinnis和Ostrom 2014)、描述该系统的现有文献以及作者对该系统的了解(Martone 2009, Shester和Micheli 2011, Micheli等人2012,2014)开发了一个概念模型b, McCay等人2014;组件被组织成资源单元,它们之间相互作用,并且是更大的资源系统的一部分,该系统受制于可以影响系统的外部力量,如市场或其他社会经济驱动力和全球环境变化。治理系统为一组参与者定义了一组规则,其中一些参与者是资源用户。这些系统及其组成部分以不同的方式相互作用,产生社会和生态结果。最后,我们指出了系统外部但影响组件及其交互的相关驱动程序和条件。
资源系统
研究合作社位于墨西哥下加利福尼亚州中部太平洋海岸的维兹卡诺半岛,该地区被称为北太平洋地区(图1)。北太平洋地区的特征是温带到亚热带,海表温度全年在12-27°C之间。该地区是由岩石礁和沙质潮下生态系统组成的马赛克,环绕着巨大海带的南缘(Macrocystis pyrifera),其中一个持续上升流区保持了较高的生物生产力(Martone 2009)。
治理
这些合作社隶属于下加利福尼亚工业和渔业区域合作社会联合会,该联合会与国家和区域渔业机构一起作为管理机构监测资源和制定管理计划。北太平洋地区的渔业合作社可以追溯到20世纪30年代末,作为墨西哥合作社运动的一种表现形式,该运动被纳入国家渔业发展政策的主流(Ponce-Díaz等人,2009年,McCay等人,2014年)。从合作社开始,获得高价值渔业,如龙虾(Panulirus中断)和鲍鱼(石决明Spp .)在邻近的渔场,法律规定只能由合作成员使用。自1992年以来,这一特别权利以20年的特许权形式出现,为包括龙虾和鲍鱼在内的一些物种提供专属开发权。
合作社持有的长期排他特许经营权是分配和管理社区使用权、退出权、排他权和管理权的例子(Schlager和Ostrom 1992年,Pomeroy等人2001年)。这些以社区为基础的获取权使合作社能够增加和执行保守的管理措施,包括但不限于龙虾渔业的捕捞组成、季节性关闭和规模限制(Vega 2001年)、特定珊瑚礁配额,以及自愿建立禁捕保护区,专门用于重建鲍鱼种群和其他渔业目标(Micheli等人,2012年,McCay等人,2014年)。合作成员和非合作成员都遵守规则,这是因为合作成员的积极和消极激励,以及在监测、执法和基础设施方面的持续投资(McCay et al. 2014)。然而,由于通常伴随管理和排他性权利而来的激励措施并不针对这些物种(Shester and Micheli 2011, Micheli et al. 2014),由于对终尾鱼缺乏特许权和相关权利,可能会阻碍该地区对这些种群的管理一个).
资源单位
北太平洋渔业的目标是岩礁食物网中各种相互作用的物种,包括捕食者,如龙虾和几种鳍鱼,以及它们的猎物和竞争对手。目前,除了龙虾和鲍鱼外,合作社还对其他一些底栖物种拥有专有权,其中包括波状裹头螺Megastraea undosa,海参Parastichopus parvimensis,红海胆Mesocentrotus franciscanus,以及红藻石花菜robustum.合作社也捕捉鳍鱼,主要是鲈鱼(Paralabrax nebulifer)、海白鱼(Caulolatilus及加州大比目鱼(Paralichthys californicus)使用渔网和捕兽夹。与底栖无脊椎动物和藻类相比,渔业合作社并不拥有鳍鱼的领土权利,因此非渔业合作社成员的渔民也可以捕捞这些物种。
演员
FEDECOOP渔场位于一片广阔沙漠的海岸边缘,自1988年起受到教科文组织生物圈保护区指定的保护(图1)。这些渔场的偏远性质,加上它们独特的制度结构和沿海岸的占领历史,导致合作社在社区中发挥着强大的作用。合作社共同为社区的许多居民(包括合作社成员和非成员)提供基础设施、社会项目和就业机会,从事涉及收获、规则执行、资源监测、海产品加工和运输的工作(Ponce-Díaz等,1998;图1)。
社会经济和生态的相互作用及其结果
加入合作社会带来社会和经济效益。鲍鱼和龙虾是主要目标,由于这些商品的需求和市场价格高,它们为合作社提供了高价值(McCay et al. 2014)。鳍鱼渔业在该地区也具有重要的经济意义,在某些情况下是合作社额外的、甚至是最重要的收入来源(Shester和Micheli 2011年)。此外,在主要目标鱼种的封闭捕捞季节,从事鳍鱼捕捞的机会为合作社成员提供了额外的收入,也为社区中的非合作成员提供了收入和自给自足的收获,他们无法获得属于合作社权限的底栖渔业目标(Shester和Micheli 2011年)。然而,尽管为合作成员和非成员渔民提供了重要的利益,但这种渔业使用的渔具类型往往有更高的副渔获率,并可能对目标种群、食物网和栖息地产生不利的长期影响(Shester and Micheli 2011, Micheli et al. 2014)一个,b).
循环分析
循环分析首先将概念模型(图1)转化为SES组件之间交互作用的图形表示,包括资源单位(如物种、渔业渔获量)、行为者(如合作成员和非合作渔民)、治理系统(如收获控制规则、捕捞努力)和社会经济因素(如收入、非合作成员的工作;我们在我们的概念模型(图1)中收集了一些主要的生物和社会经济成分,并在有符号的图表中说明了它们之间的联系(图2;(附录1)。我们将生物联系表示为目标物种或物种群之间可能的营养关系、基于收获控制规则的研究区域内物种和渔业之间的联系、以及系统的关键参与者和一些相关的社会经济成分(图2)。交互网络是根据作者对系统的经验和知识以及发表的文献中描述的关系组装的。包括对底栖动植物的采样和分类鉴定、鱼类和无脊椎动物胃内容物分析、对渔民和合作成员的采访以及住户调查(Shester 2008, Martone 2009, Morales-Zárate et al. 2011, Ramírez-Sánchez et al. 2011, Shester和Micheli 2011, McCay et al. 2014, Leslie et al. 2015)。
循环分析定性地预测相关变量(如物种丰度或渔获量)的平均变化,以响应改变其变化率的变化条件,即控制变量变化率的参数的变化。一个例子是增加一个物种死亡率的压力源。这降低了该物种的种群增长率,进而影响到该物种的数量,以及与该物种在网络中相连的其他物种的数量。一种成分水平的变化j由于一个参数的变化可以通过循环公式计算:
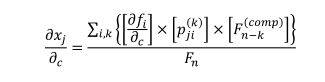 |
(1) |
在哪里c是变化的参数(如死亡率、繁殖力、捕食率);∂fi/∂c的增长率是否i变量为递增、递减(分别为正输入或负输入);p霁(k)是连接经过参数变化的变量,我,用计算其平衡值的变量,j,其中包括k变量;Fn - k(排版)是互补反馈,它缓冲或逆转通路的影响。分母表示系统的整体反馈,是衡量整个系统变化的惯性。在附录1中给出了更详细的循环分析方法和预测算法的解释。循环分析模型使用附录2中提供的代码在R中运行。
生物的链接
对于我们的资源单位,我们在循环分析中包括三个主要目标物种和物种复合体:龙虾种群、鲍鱼种群和鳍鱼种群。我们展示了社会-生态系统中的以下生物相互作用:龙虾是包括鲍鱼在内的软体动物的已知捕食者(Braje et al. 2009);鳍鱼,如海湾石斑鱼(Mycteroperca jordani)、大头鱼(Scorpaenichthys marmoratus)和羊头(Semicossyphus舰上)是龙虾(尤其是幼虾)和鲍鱼的捕食者(Braje et al. 2009;因此,鲍鱼(AS)、终尾鱼(FS)和龙虾(LS)形成了一个三营养系统:FS捕食AS和LS,而LS捕食AS。所有种群自身都有负反馈循环,代表着密度对种群增长率的依赖效应。因为这些捕食者都是多面手,而这些相互作用驱动自上而下或自下而上过程的程度在这个系统中是未知的,我们在三种不同的模型中测试了这些相互作用在网络中的影响,包括自顶向下和自底向上效应,只有自底向上效应,以及没有生物相互作用。后一个案例探讨了在决定鱼类变量动态方面,生物相互作用完全被社会经济联系所掩盖的情况。
渔业的链接
北太平洋合作渔业使用陷阱捕捉龙虾,使用水烟潜水捕捉鲍鱼,使用各种齿轮类型捕获鳍鱼,包括陷阱、集刺网和漂网(Shester和Micheli, 2011年,Micheli等人,2014年)b).与大多数渔业一样,我们假设种群和努力对渔获量有积极影响,渔获量对种群有消极影响,努力和种群相互间有消极影响(图2)。种群和努力之间的负联系考虑到种群越大,获得相同渔获量所需的努力就越低。在模型中,控制渔业的因素被转换为努力和渔获量的自阻尼项,代表模型中不包括但可调节模型组成部分的其他变量的作用(Bodini, 1988年)。尽管除了密度依赖机制外,还有几个因素可以在变量上产生一个自阻尼项(Puccia和Levins 1986),但我们没有在所有的变量中包含这个机制,因为在模型中包含所有对系统具有调节作用的变量会使模型变得难以处理。由于这些渔业都是在同一个合作体系下由相同的渔民进行的,鲍鱼和龙虾是主要的合作对象,我们将渔业之间的努力联系起来,这样龙虾和鲍鱼的努力都会负向影响鳍鱼努力的变化率,鲍鱼的努力与龙虾的努力负向相关。通过改变我们的模型结构,我们测试了包括这些联系对SES结果的影响(图2)。
除了合作社内捕鱼工作之间的联系之外,还有一些非成员也可以获得终尾鱼,因为合作社不拥有对终尾鱼的独家访问特权。我们通过将非成员努力与合作社目标的同一鳍鱼种群负相关来单独捕获非成员努力(图2)。非成员努力与非成员鳍鱼捕获量正相关,与鳍鱼种群大小负相关(图2)。
社会经济关系
合作社成员和非成员都直接从捕获的鱼类中获得收入。因此,非成员的长须鱼捕捞量正向影响非成员的收入,而长须鱼、鲍鱼和龙虾捕捞量都正向影响成员的收入(图2)。在我们的所有模型中,收入都负向影响努力。此外,为了体现合作社通过提供工作机会(例如在海鲜加工厂)对这些社区非成员的积极影响,成员收入积极影响社区的工作机会,工作积极影响非成员收入(图2)。
通过循环分析预测变化
循环公式(1)允许预测系统组件的水平如何因外力而变化(Puccia和Levins 1986)。用循环公式计算的预测可以排列在一个表中,用符号表示预期的变化方向(+、-或0)。在图3中,预测的符号用箭头代替,以便更清楚地表示。当正参数输入影响每个行变量时,表中的条目表示列变量中预期的变化。表中的每一行都表示受参数变化影响的变量。变量对给定行变量变化率变化的响应报告在列中。这些反应涉及变量(例如生物量、个体数量或金钱数量)水平的变化方向。预测通常是由正输入得到的。在负输入的情况下,预测的变化方向沿着感兴趣的行简单地倒转。
模型变量通常通过多条路径相互连接。如果这些途径有相反的效果,该模型就会产生模棱两可的预测。在这些情况下,模型预测是不确定的,并且预测表中的+或-符号被问号(?)所取代。为了解决这些不明确的预测,我们使用了一个程序,随机分配数值给社区矩阵的系数(即,有符号有向图中链接的系数)。链接的值由例程在间隔(106- 1).执行此程序nxnX 100倍,其中n是模型中变量的数量。因此,我们为每个模型创建了团体矩阵(n= 14;总跑步数= 19,600)。然后,可以对群落矩阵进行倒序,以了解变量如何通过间接途径直接影响彼此(Bender et al. 1984, Wootton 2002, Montoya et al. 2009)。系数(cij-1)表示变量的整体效应j在变量我由于它直接链接到变量我(例如,捕食,捕获),以及通过该变量的所有可能的间接途径j被连接到我通过中间组件。因此,一个扰动对变量的净效应(直接和间接效应的总和)j在变量我由团体矩阵逆的元素给出。
社区的矩阵啊必须有一个非零行列式并且必须有一个逆矩阵(啊)-1.的nxn为每个模型创建x 100个矩阵,只保留那些满足李雅普诺夫稳定性条件的矩阵并将其倒置。然后从倒立矩阵得到每个模型的预测总表。在这个表中,每个预测都是根据反向稳定矩阵数组中正号、负号或零号的百分比来确定的(附录1,表S2)。我们定义了一组规则,将从循环分析运行获得的案例百分比转换为整体预测矩阵中的符号(Puccia和Levins 1986)。具体来说:-表示0-25%的关系是积极的;-表示25-40%的关系是积极的;0*表示40-60%的积极变化关系;?+表示60-75%的联系是积极的;a +表示75-100%的信号为阳性。这是基于Puccia的3:1比率规则(Puccia and Levins 1986)。 Note that 0* are not real zeros but a neutral result that occurs when matrices have large numbers of opposite signs for a given variable’s response. In fact, when multiple pathways have opposite effects to the same variable, positive and negative effects tend to compensate each other and the net result may be zero (no variation) or, more likely, a small change, which can be reasonably considered negligible.
模型结构和场景测试
我们通过进行两种不同类型的分析来检查系统动力学和行为。首先,我们探讨了关于模型结构的不同假设的影响,特别是变量之间的联系被包括或排除(五个模型结构)。其次,我们研究了系统对摄动的响应(考虑7种不同的摄动场景)。
为了检验模式结果对所包括的具体联系的敏感性,我们改变了模式结构,包括或删除了不同的生物和(或)渔业联系。模型1-5(图2;附录1,表S2)包括所有的生物联系,在其中,捕食者和猎物的影响都被表示出来,但每个模型不同的捕鱼努力之间的关系,包括或删除鲍鱼和龙虾渔业、龙虾和鳍鱼渔业、鲍鱼和鳍鱼渔业之间的假设联系。
我们还改变了核心生物结构,以检验不同物种如何通过消费者-资源交互作用相互影响的假设的结果的稳健性。在第二组模型中,我们删除了所有的生物联系,并改变了捕鱼努力之间的关系,如上面的模型1-5所述。在第三组模型中,我们包含了生物联系,但仅以猎物对捕食者产生的有益影响的形式存在,即资源对消费者产生的自下而上的影响。在所有这些情况下,模型得出的矩阵行列式为零,无法得到任何预测。这意味着,要产生有意义的预测,模型需要完整的生物相互作用。因此,我们只提出和讨论模型1-5的结果(图3;附录1,表S2)。
我们研究了单个变量对系统参数变化的响应,方法如下:(a)每个模型的预测表,以探索生态和社会经济组成部分之间的关系,以及三种渔业内部的关系;以及(b)七个与特定外部扰动和管理行动相关的场景,以检查感兴趣的变量的响应,包括生物变量(种群丰度)和社会经济变量(工作、收入)。
环境扰动
疾病、缺氧和气候变化是海洋生态系统变化的主要beplay竞技驱动因素,并与许多沿海渔业渔业物种死亡率的增加有关(Defeo和Castilla 2012, Micheli等,2012)。在情景1中,我们通过鲍鱼生长速率的下降来模拟气候或其他人为影响的外部强迫。缺氧、频繁或极端的厄尔尼诺-南涛振荡(ENSO)事件、疾病或有害的藻华是观察到的鲍鱼数量下降的基础,并可能通过增加鲍鱼死亡率导致进一步下降(Morales-Bojórquez等,2008年,Micheli等,2012年)。在情景2中,我们将疾病或海洋酸化对龙虾种群的影响建模为对龙虾种群的负面输入,反映了这些外部驱动因素导致龙虾死亡率增加或龙虾生长和繁殖减少。关于海洋酸化对甲壳类动物影响的研究表明,由于血淋巴中[H+]的增加和向组织输送氧气的减少,可能对生长产生负面影响(参见Whiteley 2011年的综述)。尽管疾病没有影响到该系统中的龙虾种群,但它是其他龙虾渔业的主要问题(例如,Steneck等人,2011年)。
社会经济驱动程序
我们研究了市场化举措的效果,如现有的海洋管理委员会(MSC)生态标签或拟议的全系统认证计划,其实施目标是增加渔业合作社的收入(Micheli等人,2014年)一个).我们在情景3中将这些驱动因素建模为合作社成员收入的积极投入,尽管在维兹卡诺合作社的情况下,尽管生态标签允许在美国市场获得更高的价格,但它主要是赋权的来源,并帮助合作社维持其特许权(Pérez-Ramírez等,2012年)一个,b).还为该区域提出了其他倡议,目的是增加非合作成员的就业机会,例如鲍鱼珍珠文化和生态旅游。在情景4中,我们通过为非成员提供渔业替代方案的合作社提供的这些机会,测试了变化对非成员收入的影响。
渔业管理措施
尽管尚未实施,但减少捕捞努力和逐步淘汰固定刺网已被强调为减少环境影响和提高这些合作渔业的长期可持续性的可能选择(Peckham等人,2007年,Shester和Micheli 2011年,Micheli等人,2014年b).在场景5中,我们研究了这些管理行为如何影响系统中的其他变量,使用一个消极的输入来影响合作成员的最终努力。在场景6中,我们检查了对系统上非成员finfish努力的控制的影响。最后,在情景7中,我们考察了通过政府、私人基金会或非政府组织提供的补贴而增加的就业机会的影响。
敏感性分析
为了检验模型结构是否影响结果,我们比较了五种模型的预测表,看看它们之间是否一致。在一系列两两比较中,我们将每个模型的每个预测矩阵与所有其他矩阵进行了比较,并确定了每个预测矩阵与所有其他矩阵不同的情况的数量(附录1,表S3)。
结果
模型预测:社会生态系统(SES)动力学
循环分析表明,输入变量(例如股票增长率、成员收入变化率)的变化可能会影响系统中其他变量的方向,而这些方向往往无法根据已知(或假设)的方向关系进行预测。例如(见图3),鲍鱼生长速度的增加(对AS的正向输入)被预测会减少龙虾的数量,这与被捕食物种应该积极影响捕食者数量的概念相反。这是因为一个变量对任何其他变量的影响是由系统中的其他变量通过间接效应来中介的。
另一个令人惊讶的结果是捕食者的变化对猎物没有影响。捕食者和被捕食者之间的相关性取决于被干扰的种群,因为在整个SES中有反馈和补偿。例如,当finfish种群(FS)的增长率增加时,FS本身被预测会增加,但鲍鱼种群(AS)没有变化(图3)。这一结果可以解释为FS捕食AS,同时也捕食LS,而LS又捕食AS。所以FS对AS的影响是积极和消极的,因为这个物种也以AS的捕食者为食。然而,SES的复杂性增加了连接FS和AS的路径的多样性。循环分析表明FS与AS之间有8条路径:4条带负号的路径和4条带正号的路径(链接符号的乘积产生路径的整体符号;见附录1,表S1)。因此,我们的模拟得到了预期AS将增加和预期AS将减少的相同比例的矩阵(50%),从而产生了补偿效应和接近于零的变化幅度。
FS的正输入导致龙虾种群(LS)减少。当正向输入影响FS的生长速度时,LS所经历的这种负面影响可能是由于对LS的直接捕食和对猎物的捕食。然而,在像我们在图2中描述的这样的复杂系统中,一个变量对另一个变量的任何影响都是由多个途径介导的,因此必须检查所有途径才能理解其影响。
9条路径连接FS和LS(图2;附录1,表S1)。5种对LS有负影响,4种对LS有正影响。尽管只有一条额外的路径表明FS对LS的负面影响,但数值模拟结果显示,只有8%的矩阵预测当正向输入影响FS时LS水平会增加。但91%的矩阵预测LS种群在FS正输入后会减少。这是令人惊讶的,考虑到携带相反影响的准平衡数量的路径,我们应该期望模拟的结果更加平衡。我们所期望的路径数量与我们从模拟中获得的结果之间的差异可能取决于以下几点:随机系数取在10范围内6- 1,但较长路径上的链接值的乘积比较短路径上的链接值的乘积要小;因此后者对循环分析的最终结果贡献更大。如果我们考虑FS到LS的路径(附录1,表S1),两条最短路径都有负号,我们可以理解为什么尽管相反路径之间存在数值上的准平衡,但LS对FS增长率增加的响应是负的。在前面的例子中(输入到FS和影响到AS),相反,两条较短的路径(附录1,表S1)有相反的符号。FS和LS之间的这种关系与已知的捕食者-被捕食者关系的预期是一致的,一些当地渔业的目标鱼种(如羊头鱼、美国现象,)捕食龙虾(即FS增加对LS的预期负面影响)。
尽管认为FS和LS作为捕食者会对AS产生负面影响,但当AS受到干扰时,AS与FS之间呈现正相关关系,而AS与LS之间无相关关系或负相关关系。当输入通过龙虾存量(LS)进入系统时,LS与As呈负相关,而这两个变量与FS之间不存在相关性,在所有5个模型中,预测FS的丰度不会发生变化。
预测表可作为检测扰动入口点的诊断工具。例如,只有当扰动通过鲍鱼和龙虾渔业以输入的形式进入系统时,鲍鱼种群(AS)才会发生变化。唯一的例外是模型4(附录1,表S2),在该模型中,合作社(ME)成员对finfish的投入预计会影响鲍鱼种群。根据这些结果,AS的任何变化都可能与鲍鱼或龙虾渔业的变化有关,但与系统的其他变量(即经济变量)无关。
审查渔业变量(渔获量、努力量和种群)之间的关系也可以提供对特定渔业动态的洞察。例如,包含变量AS、AE和AC之间关系的3 × 3子矩阵(鲍鱼存量、捕捞努力和捕获量;可以检查图3)以收集有关鲍鱼渔业各方面的信息。正如预期的那样,鲍鱼储量与努力程度呈负相关:当努力程度增加时,储量减少,反之亦然。努力和渔获量在一个方向上呈正相关,但在另一个方向上呈负相关:随着努力增加,渔获量增加,但随着渔获量增加,预计努力将减少。而渔获量与渔获量呈中性关系,表明努力是关键控制变量。这个子矩阵在我们研究的所有五个模型中都是相同的(附录1,表S2),表明基于这个子矩阵的结论对模型结构的变化具有鲁棒性。
预测表还可以指出哪些变量最容易受到来自环境变化或管理干预的外部扰动的影响。例如,成员收入(MI)比非成员收入(NI)对参数变化表现出更大的惯性(图3)。在模型1中,MI对外部输入的14个可能响应中有8个为空(图3),而在其他模型中这种模式仅发生轻微变化(0的数量在6到8之间变化;附录1,表S2)。只有当输入通过AE、LS和ME以五种模型的不同组合方式进入系统时,非成员收入才保持不变。因此,与非成员收入相比,合作成员收入似乎不太容易受到扰动的影响,但也被预测对管理干预的反应较小。
预测自然扰动和管理行动的结果
目标分类群的崩溃(场景1和2)
疾病和气候变化是目标物种beplay竞技死亡率增加和渔业崩溃的主要驱动因素。在情景1中,我们模拟了一种疾病、极端ENSO、有害的藻华或缺氧事件对鲍鱼的影响(Shepherd等人,1998年,Morales-Bojórquez等人,2008年,Micheli等人,2012年)作为鲍鱼种群的负面输入(as;图4).如果没有有效的努力控制,鲍鱼种群的减少伴随着努力的增加,而渔获量没有变化。然而,会员和非会员的收入预计会减少,因此鲍鱼死亡率的增加会导致当地社区的整体经济损失。从生物学角度看,鲍鱼种群的减少伴随着鳍鱼种群(FS)的减少和龙虾种群(LS;图4)。有趣的是,预测成员(ME)和非成员(NE)的捕尾鱼努力都将增加,但两组(MC, NC)的捕尾鱼量都将下降(图4)。在所有模型下,工作机会(JO)都不受影响。
在情景2中,我们模拟了一种疾病对龙虾种群的负面影响(Steneck et al. 2011)。对龙虾种群的负面输入预计将导致鲍鱼种群的增加,而对鳍鱼种群没有影响(图2),假设疾病不会同时影响鲍鱼和鳍鱼种群。这些模型揭示了系统对龙虾种群扰动的巨大惯性,因为所有其他渔业和社会经济变量都没有发生变化(图4)。这些模拟强调了系统对鲍鱼的扰动比龙虾的扰动更敏感,这从场景1比场景2更多的负面结果可以看出(图4)。
情景1和2的结果对于系统中相互作用的表示相对稳健,因为通过努力变量删除渔业之间的联系不会改变变量的方向响应(附录1,表S2)。
基于市场的干预措施(情景3和情景4)
在情景3中,我们检验了生态标签的效果,它代表了对会员收入(MI)的积极投入。这是唯一能对生物和经济变量以及两种用户群体产生积极结果预测的场景。预计成员和非成员的收入和就业机会都将增加,所有渔业的捕捞努力预计将减少,鲍鱼和龙虾的数量预计将保持稳定,而鳍鱼的数量预计将增加(图4)。因此,通过市场激励措施提高成员收入预计将对整体经济产生有利影响,对稳定(AS和LS保持不变)或增加(FS)的数量产生积极影响。
在情景4中,非成员收入通过旅游业发展的机会或与渔业无关的其他收入机会而增加。在这种情况下,福利只会对非会员产生,而会员的收入预计将保持不变。这种干预不会对鲍鱼渔业产生影响,as、AE或AC没有变化。由于与经济替代品相关的捕捞努力减少,鳍鱼的数量将增加,而龙虾的数量预计将减少,因为它们的捕食者数量增加。因此,在这种情况下,主要目标股票和整体经济预计不会受益。
渔业管理行动(方案5、6、7)
为减少成员在鳍鱼渔业(ME)的努力而采取的管理措施预计将导致所有用户群体的负面社会经济结果(情景5;图4).预计会员和非会员的收入和工作都将减少。因此,从经济上讲,这种情况的后果是最不利的。成员的终末鱼捕获量(MC)呈下降趋势,而非成员的终末鱼捕获量(NC)呈上升趋势。对鲍鱼(AS)和龙虾(LS)的预测结果不同:预计鲍鱼(AS)的数量将下降,而预计AS不会发生变化。有趣的是,尽管鲍鱼数量和捕获量都在增加,AS却没有变化。这突出了使用线性因果关系标准预测结果的难度,该标准是通过关注单个渔业中种群、努力和渔获量之间的独立相互作用而制定的。
非成员finfish努力的减少(场景6)也预计将导致非成员的经济损失,而成员的收入没有提高。非会员收入预计将减少,可能是由于努力减少导致捕捞减少。鱼类资源增加导致龙虾资源减少,而鲍鱼资源则不受影响,鲍鱼渔业似乎对这种投入不敏感。随着鳍鱼数量的增加,渔业内部的反馈导致了不断的捕捞,即使减少了努力。这并不适用于非成员,因为捕获和努力都减少了。
增加工作机会的变化率(设想7),例如通过外部补贴或投资,只会产生有限的有益效果。与前一种情况(情景6)一样,非会员收入预计将增加,而会员收入将保持稳定。鲍鱼渔业似乎没有受到这种干预的影响,而龙虾渔业则有负面影响(图4)。
我们进一步改变模型结构,以检查可能影响系统的其他管理干预的潜在结果。具体来说,在我们的模型1-5中,我们代表了应对渔获量和种群变化的努力的更一般情况,并没有考虑可能影响这种关系的监管控制,假设监管控制可能并不总是有效。从1960年至2010年(Vega 2001年)下加利福尼亚龙虾的捕捞趋势和努力数据可以看出,历史记录中有两个阶段。在第一个时期(从1970年到大约。1990年)努力增加,而渔获量基本保持不变;在第二个阶段(1995 - 2004年),渔获量随着努力的持续而增加。从我们的模型预测表明,只有当我们模拟龙虾库存的负面输入时,例如死亡率增加或招募减少,努力的增加可能伴随着恒定的捕获。反映第二种模式的结果,即持续的努力伴随着渔获量的增加,需要对龙虾渔获量作出积极的投入,这可能通过例如实施齿轮更换来反映渔获率的提高。然而,在1995-2004年期间,FEDECOOP合作社通过保持陷阱的数量和捕鱼季节的长度不变,努力保持不变(Vega, 2001年)。为了检验龙虾捕捞努力的有效控制如何影响结果,我们在其他模型中引入政府机构作为龙虾捕捞努力的控制变量(附录1,表S4)。引入政府机构(GA)作为LE的外部控制,没有自阻尼,也没有与系统其余部分的其他链接,因此GA仅对龙虾的努力作出响应和作用(附录1,图S1)。 Interestingly, the table of predictions indicates that the presence of this variable makes LE resistant to all parameter changes except for input on GA itself (Appendix 1, Table S4). Models predict that increased catch can be obtained only with a positive parameter change on lobster catch, as in the case without control over effort (Fig. 3). In this latter case, a null value represents a true zero response and not compensation due to opposite forces. This zero response is typical of variables that are connected to a satellite variable (Levins 1974, Puccia and Levins 1986). Government agency is a satellite variable because it is connected to the system only through its linkage with LE and taken in isolation represents a system with zero feedback.
敏感性分析
在考虑的5个模型中,预测征兆频率的最大差异出现在模型2和模型3之间,在所有两两比较中有28%的差异(附录1,表S3)。平均而言,五种模型之间的比较在20%的病例中产生了不同的迹象。然而,如果符号的倾向(即?+和?-之类的预测)被认为是真实的符号(即?+变成+和?-变成-),差异就不那么明显了:模型2和3仍然是差异最大的,但只有17%的比较得出不同的预测,模型之间的平均差异为12%。因此,模型结构可以影响结果,但在一小部分模拟中得到了不同的预测,这表明结果可以被认为对我们所测试的模型结构的变化是相对稳健的。
讨论
定性建模方法为了解SES中可能的行为和对干预措施的响应提供了工具(Dambacher等人,2007年,Carey等人,2014年),循环分析在墨西哥下加利福尼亚州维兹卡诺地区沿海小规模渔业的应用就是一个例子。这种方法可以对管理行动、新政策或环境驱动因素的可能结果产生预测和假设,并可以突出需要研究的关键环节,以便更好地理解复杂SES的动力学。应用于下加利福尼亚海岸SES的循环分析表明,生物和社会经济成分之间的复杂反馈是如何由于外部扰动而导致反直觉和意想不到的结果的。总的来说,我们的研究结果表明,当管理的目标是同时改善不同用户群体的环境条件和生计时,应仔细考虑管理行动和新政策的可能权衡和级联效应(Levin等,2009年,Carey等,2014年,Micheli等,2014年)一个).我们的结果与应用SES框架的下加利福尼亚小规模渔业的更广泛的分析一致,该框架强调了在实现生态和社会可持续性方面的权衡,以及下加利福尼亚半岛不同地理区域之间的高变异性(例如,Leslie et al. 2015)。
循环分析有助于突出生态系统的哪些组成部分可能更容易受到扰动的影响,以及哪些干预措施可能比其他干预措施对生态系统的不同组成部分和用户群体更有益。我们对未来环境或管理变化的可能情景的分析表明,在我们对系统组成部分如何相互作用的假设下,龙虾种群预计是最脆弱的,而在大多数情景下,鳍鱼种群预计将受益。鲍鱼种群似乎通常是绝缘的,根据大多数情况,对其他成分的变化表现出相对较高的抵抗力。然而,鲍鱼的大量死亡(设想1)预计将对不同用户群体的捕捞和收入产生负面影响,总体上的负面后果比疾病或环境条件导致的龙虾死亡更多。换句话说,鲍鱼种群对其他变量的变化相当不敏感,但鲍鱼的变化率的变化很可能会影响整个系统。我们的模型还显示了龙虾和鳍鱼种群之间的负相关。这种负相关表明,试图恢复两个种群中的一个或提高其增长率将对另一个产生负面影响,除非干预措施针对少数变量(即AC、LS、LC、ME、MI)。在管理干预之前,需要对这些假设进行检验和仔细考虑。
模型产生了关于不同干预或扰动可能产生的社会经济结果的具体预测,并突出哪些管理干预可能对当地经济最有利或最不利。只有情景3,生态标签,对所有三个社会经济变量,即成员和非成员的收入和工作,显示出积极的影响。其他六个情景对系统的经济产生的结果不那么积极,其中大多数预测都是负面的(例如,大规模死亡影响鲍鱼种群,这是该系统中最宝贵的资源,以及合作社内捕捞鳍鱼的努力减少)或没有显示一致的变化(例如,大规模死亡影响龙虾,合作社对鲍鱼拥有独家访问权)。因此,在这一系统中,在考虑的管理干预措施中,生态标签预期具有最大的效益,而减少合作成员的捕渔努力是最有害的。此外,鲍鱼大规模死亡事件的经济影响预计将大于龙虾的情况。
在该区域进行的其他研究记录了渔民和渔业在面对环境条件变化时的不同表现。例如,Finkbeiner(2014)发现,渔业活动的多样化对于降低风险和稳定收入很重要,但在有利条件下专攻高价值物种的能力会带来财富积累。因此,在环境条件不断变化的情况下灵活地调整各种捕鱼战略对小型渔业合作社的适应能力是很重要的。对下加利福尼亚州和其他地区的SES的进一步研究应该考虑到这些对变化的动态响应,以及不同的治理框架和市场如何促进或限制适应。
模型还强调了哪些用户群体可能更容易受到外部冲击的影响。在这个SES中,非成员收入比成员收入对环境变化或在这些场景中考虑的管理决策更为敏感(图4;附录1,表S2)。有趣的是,当系统因影响非成员的输入而受到干扰时,如鳍鱼存量、非成员收入、非成员对鳍鱼的努力、非成员渔获量和工作机会,成员收入就会显示出对变化的抵制。如果这些结论经得起进一步审查,它们将表明管理决策可能如何影响这些社区,因为成本和收益可能分配不均。这一点尤其重要,因为资源获取方面的不平等、过程的不合法和社会资本的损失会影响人们遵守法规的方式,如果忽视,可能会导致意想不到的后果,如偷猎增加和物种数量下降(McClanahan et al. 2009)。最终,这会导致贫困陷阱,影响社会经济地位的适应能力(Cinner 2011)。一个重要的下一步将是对这个和其他沿海经济社会系统的社会动态和干预可能产生的意外后果进行深入分析和建模(例如,Finkbeiner 2014)。重要的是要认识到,如果没有从整个系统的角度解决问题,治理方法在一个SES维度中解决问题可能会在其他维度中引发意想不到的后果。对该系统和其他系统中SES的全面、综合的理解将使可持续发展科学能够更充分地为可持续发展实践提供信息(Leslie等人,2015年)。
循环分析还可以揭示理想的生态和社会结果之间的重要潜在权衡,并有助于确定哪些干预措施可能导致潜在的双赢结果。例如,在本案例研究中,管理干预,如额外的finfish渔业监管,预计会产生负面的经济影响,而通过生态旅游或补贴创造新的就业机会可能会对合作成员和非成员产生混合影响。市场干预,如生态标签,预计会产生总体积极的经济效应。此外,在大多数模型结构下,这种积极的经济影响与缺乏负面的生物影响相关,这表明这种基于市场的干预造成的权衡在所有被考虑的模型中最少。此外,在这种情况下,利益被分配到多个利益相关者群体中,其中成员和非成员都有望发展壮大。然而,我们警告在没有进一步建模或对至少一组核心变量的经验支持的情况下应用循环分析的结果。例如,我们假设生态标签会自动增加成员收入的变化率,但这一假设并不适用于所有系统。虽然生态标签可以影响市场价格和增加收入,并确实为该地区的FEDECOOP合作成员和非成员带来了有记录的好处(Pérez-Ramírez等,2012b),这高度依赖于市场准入、基础设施和处理能力(Pérez-Ramírez等,2012一个)
循环分析可以确定哪些途径可能对结果有更大的影响,突出未来研究和分析中需要检查的潜在重要关系。正如我们的结果所描述的,循环分析可以预测预期的和意外的结果。例如,在这个案例中,循环分析预测鱼类资源丰富度(FS)的增加对其猎物龙虾(LS)的生长速率有负面影响,但对鲍鱼(AS)的猎物没有影响。正如在任何复杂系统的网络分析中一样,多个路径调解了一个变量对另一个变量的影响,因此必须检查完整的路径集以理解其影响。在每一种情况下(从FS到LS和从FS到AS的链接),都有多个路径具有相反的影响,即消极和积极。因此,如果只有路径的数量影响循环分析的结果,我们预计模拟的结果将导致FS对AS和LS的中性效应。我们基于路径数量所期望的结果与我们从模拟中获得的结果之间的差异很可能取决于这样一个事实:从随机分布中提取的链接值的乘积可以在较长的路径中产生较小的数字,而较短的路径往往会驱动循环分析的最终结果。在上面讨论的例子中,从FS到LS的最短路径都是负的,但从FS到AS的最短路径都是正的和负的。
这个案例研究举例说明了SES应用程序中循环分析的潜力,并强调了这种方法的其余弱点和注意事项。在循环分析中,与其他建模方法类似,预测强烈依赖于对SES相关组件的具体假设、这些组件之间联系的性质和网络的整体结构。不确定性的一个主要来源与我们准确表示变量之间关系的能力仍然有限有关。尽管我们对维兹卡诺渔业社会生态系统的描述和关于相互作用的性质和方向的决定是基于对这个系统和其他类似系统的现有知识,但仍然存在高度的不确定性。例如,我们在几个模拟中观察到的对龙虾种群的负面影响很可能是由于假设某些鳍鱼类群(例如,被当地渔业瞄准的羊头鱼)的捕食控制了龙虾种群。尽管羊头是龙虾的主要捕食者,而且在南加州的海带森林中已经证明了羊头对底栖无脊椎动物种群的控制(Cowen 1983),但没有经验证据表明在下加利福尼亚海岸对龙虾种群的自上而下的捕食控制。
提高预测的可靠性可以通过设计替代网络结构和评估预测对这些联系的不同描述的稳健性来实现。这可以帮助确定哪些结构差异是重要的。例如鲍鱼渔业子系统(图3,标记为AS、AE、AC的列)似乎对环境变化具有相当的抵抗力。在我们分析的5个备选网络结构中,与该子系统相关的42个预测中,22个或更多的预测对参数变化没有响应(附录1,表S2)。这个稳健的结果可能是所有模型共有的核心结构的影响,在这个结构上添加或删除一些链接并不会改变预测。因此,特别重要的是要仔细构建模型的核心结构,集成所有可用的信息来源和来自不同涉众的输入。事实上,循环分析的一个重要好处是,它为集成不同类型的信息提供了一个框架,从而为参与式模型的构建提供了一个让涉众参与的机会(Anthony et al. 2013)。对于定性和定量建模方法,模型开发将受益于实施参与式框架,其中包括来自不同用户的不同类型的系统知识,并对其进行测试,以确定哪些变量和联系与结果最相关(Essington等人,2016,Stier等人,2016)。
我们的结果,除了在不同情况下的特定预测外,强调了相关模式取决于网络结构和摄动的入口点。例如,当生物压力进入系统(情景1和情景2)时,鲍鱼(AS)和龙虾(LS)种群之间是负相关的。然而,当压力扰动影响非生物变量(努力、收入、工作机会,情景2到7)时,这两个种群之间就显得不相关了。从现场数据的相关模式可以用来检测系统中的扰动入口点,这突出了回路分析作为诊断工具的潜力。
循环分析已用于渔业背景下的其他几个场景分析案例(Anthony等人,2013年,Espinoza-Tenorio等人,2013年,Carey等人,2014年,Dambacher等人,2015年)。然而,这种技术存在一些限制,必须在管理上下文中仔细处理。Justus(2006)概述了循环分析的结果与变量平衡水平的变化有关,但实际系统通常不处于平衡状态,非线性常常表征变量的动力学。尽管之前的研究(Lane and Collins 1985, Bodini 1988, 2000)已经提供了证据,证明循环分析可以通过考虑变量平均值的变化而应用于动态系统,但动态变化仍然是定性建模技术的主要挑战。循环分析的另一个限制是预测的影响可能显现的时间尺度。例如,对于SES的某些组件来说,变化可以在很长一段时间内逐渐发生,而其他组件则可能经历突然的变化。属于不同领域的变量可能表现出非常不同的动态,因此必须仔细考虑时间尺度,并结合定性分析和定量动态模型直接处理。
与循环分析相比,其他定性建模方法可以提供一些优势。例如,模糊认知图(fcm)通过变量之间关系的半量化明确了链接的大小(Özesmi和Özesmi 2004, Kok 2009)。这种方法解决了上面讨论的对比路径的净效应的模糊性。FCM相对于循环分析的另一个优点是,它可以对多个同时发生的扰动进行预测。然而,构建fcm的过程要求专家使用模糊条件(IF-THEN)语句描述网络的结构和相互连接。这样的表述是这样的:“如果变量A的水平高,变量B的水平就低。”这意味着专家从观察系统得到的变量之间的相关性中定义关系(Stylios和Groumpos 1999)。我们的结果强调了相关模式依赖于网络结构和摄动的入口点。例如,当生物压力通过大量死亡率进入系统时(情景1和情景2),鲍鱼(AS)和龙虾(LS)种群之间是负相关的。然而,当扰动通过非生物变量影响系统时(例如,努力、收入和工作机会的变化,情景2-7),这两个种群之间就显得不相关了。由此可见,在相互关系的基础上定义相互作用可能具有误导性(Levins and Puccia 1988)。
循环分析和fcm都允许预测复杂系统中某个变量受到扰动后的变量水平的变化。然而,循环分析的一个优点是它模拟了扰动对变量变化率的影响,而不依赖于变量的初始状态。例如,在情景1中,我们调查了气候变化导致鲍鱼死亡率增加的影响。beplay竞技根据现有数据(Morales-Bojórquez et al. 2008, Micheli et al. 2012),尽管我们可以预计热浪和/或缺氧会增加鲍鱼种群的死亡率,但更难预测由此产生的丰度。相反,在fcm中,扰动被模拟为一个或多个变量(即总体的丰度)初始状态的变化,但变量水平的变化很难预测(Levins和Puccia 1988)。因此,在我们的例子中,循环分析更合适,因为它允许基于对变量变化率(负的、中性的或正的)方向的知识的干扰的合并。考虑到每种方法的优点和缺点,Ramsey和Veltman(2005)表明,有很大的潜力进行富有成效的集成。
其他定性建模方法包括因果循环图(CLD;Richardson 1986)和贝叶斯信念网络(BBN;Borsuk等人。2004;pollo等人。2007)。因果循环图通过在链接极性的基础上逻辑地重构变量之间的因果链来进行预测。然而,通过使用链接极性识别反馈效应来预测复杂网络的行为是困难的,并且可能导致对效应的误导性解释,正如Richardson(1986)先前强调的那样。同样,指定BBN所要求的相关条件概率可能是一个费时费力的过程(Ticehurst et al. 2007)。此外,通过循环网络结构包含反馈需要依赖广泛参数化的动态时间显式bbn。因此,bbn通常不包括生态系统共有的反馈(Marcot et al. 2001)。与fcm类似,将bbn与循环分析相结合在改进预测和模型验证方面具有很大潜力(Melbourne-Thomas等人,2012年,Anthony等人,2013年)。 However it must be emphasized that these applications of BBNs are based on the signs derived from the analysis of the qualitative models. As such, their outcomes are contingent on the assumptions and limitations of the signed diagraph models.
结论
以生态系统为基础的方法管理渔业需要将重点从单一渔业和部门转移到综合战略,其中管理考虑到多个渔业、多个物种、当地社区的多个方面及其联系。只有从整体上看待社会经济系统,我们才能开始预测多种人类活动、管理干预和环境冲击的后果。定性建模方法允许管理人员、利益相关者和科学家检查一组可能的干预措施中哪一种对系统中其他生物和社会经济成分的影响最大,以及哪一种对变化的抵抗力更强。此外,模型及其结果可以帮助确定关键组成部分和联系,并确定数据收集和管理行动的优先次序。然而,包括环分析在内的定性模型具有很高的不确定性,模型扰动的结果取决于结构联系的假设。因此,定性模型应该用经验数据进行检验,并与定量动态模型相结合,以增加模型预测的确定性。尽管存在局限性,但包括整个社会经济体系的联系的定性模型是解决生态完整性、代际机会和经济效率这三个可持续发展范式关键方面的全面、全系统方法的组成部分。
致谢
这项工作得到了美国国家科学基金会的资助(OCE-0410439和DEB-1212124)和帕卡德基金会的保护和科学计划的资助。我们感谢B. McCay、G. Shester、S. Lluch、G. Ponce、S. Guzman del Proo、E. Serviere、D. Aguilar、M. Valenzuela以及“Progreso”和“Punta Abreojos”合作社的几位成员的建议和支持。感谢Stefania Favilla(意大利Modena-Reggio Emilia大学),她开发了第一个版本的算法,通过模拟计算环分析预测。两位匿名审稿人和Kasper Kok博士对本文的改进做出了很大贡献。
文献引用
安东尼,K. R. N, J. M.丹巴赫尔,T.沃尔什,R. A.比登。2013。一个了解累积影响、支持环境决策和为大堡礁世界遗产区基于复原力的管理提供信息的框架.澳大利亚海洋科学研究所,汤斯维尔,澳大利亚;CSIRO、霍巴特、澳大利亚;NERP决策中心,墨尔本大学和大堡礁海洋公园管理局,汤斯维尔,澳大利亚。(在线)网址:https://www.environment.gov.au/system/files/resources/2910cf7e-30fc-466f-a6c1-0e27aa618d05/files/framework-resilience-based-management_0.pdf
本德,t.j.凯斯,m.e.吉尔平,1984。群落生态学中的扰动实验:理论与实践。生态65:1-13。http://dx.doi.org/10.2307/1939452
伯克,f . 2006。从社区资源管理到复杂系统:规模问题和海洋公地。生态和社会11(1): 45。http://dx.doi.org/10.5751/ES-01431-110145
伯克,f . 2011。恢复统一:海洋社会-生态系统的概念。页面9-28在R. E. Ommer, R. I. Perry, K. Cochrane和P. Cury,编辑。世界渔业:社会生态分析.著名英国牛津。http://dx.doi.org/10.1002/9781444392241.ch2
Berkes, F.和C. Folke, 1998。连接社会学和生态系统:建立复原力的管理实践和社会机制。剑桥大学出版社,美国纽约。
Bodini, a . 1988。通过有符号的有向图表示生态系统结构。模型重建、定性预测与管理:以淡水生态系统为例。Oikos83:93 - 106。http://dx.doi.org/10.2307/3546550
Bodini, a . 2000。重建营养相互作用作为理解和管理生态系统的工具:在浅富营养化湖泊的应用。加拿大渔业和水产科学杂志57:1999 - 2009。http://dx.doi.org/10.1139/f00-153
Borsuk, M. E., C. A. Stow和K. H. Reckhow. 2004。富营养化贝叶斯网络模型的综合、预测和不确定性分析。生态模型173:219 - 239。http://dx.doi.org/10.1016/j.ecolmodel.2003.08.020
布曼斯,R.和R.科斯坦扎,2007。多尺度综合地球系统模型(MIMES):生态系统服务的动力学、建模和评估。104 - 107页全球水系统研究报告中的GWSP问题.联邦教育和研究部,全球水系统项目,德国波恩。(在线)网址:https://www.pdx.edu/sites/www.pdx.edu.sustainability/files/Boumans_Costanza_GWSP%20Chapter_2007.pdf
布莱杰,T. J., J. M.厄兰森,T. C.里克,P. K.戴顿,M. B. A.哈奇,2009。从过去到现在的捕鱼:加利福尼亚海峡群岛红鲍鱼渔业的连续性和恢复力。生态应用程序19(4): 906 - 919。http://dx.doi.org/10.1890/08-0135.1
凯里,m.p., P. S.莱文,H.汤森,T. J.米内洛,G. R.萨顿,T.弗朗西斯,C. J.哈维,J. E.托夫特,K. K.阿科马,J. L.伯克,c - k。Kim, A. Guerry, M. Plummer, G. Spiridonov和M. Ruckelshaus. 2014。用定性的联系来描述沿海食物网,以弥合基于生态系统的管理理论和实践之间的差距。国际内燃机学会海洋科学杂志71(3): 713 - 724。http://dx.doi.org/10.1093/icesjms/fst012
辛纳,2011。珊瑚礁渔业中的社会生态陷阱。全球环境变化21(3): 835 - 839。http://dx.doi.org/10.1016/j.gloenvcha.2011.04.012
C.科斯特洛,S.盖恩斯和J.林汉姆,2008。捕捞份额能防止渔业崩溃吗?科学321:1678 - 1681。http://dx.doi.org/10.1126/science.1159478
Cowen, R. K. 1983。羊头的作用(Semicossyphus舰上)捕食红海胆(Strongylocentrotus franciscanus)群体:实验分析。环境科学58(2): 249 - 255。http://dx.doi.org/10.1007/bf00399225
考克斯,M., G.阿诺德,S.维拉马约Tómas。2010.以社区为基础的自然资源管理设计原则审查。生态和社会15(4): 38。(在线)网址://www.dpl-cld.com/vol15/iss4/art38/
丹巴赫,J. M., D. T.布鲁尔,D. M.丹尼斯,M.麦金太尔,S.福勒。2007。金矿对Lihir岛社会经济系统和礁边鱼类群落影响的定性模拟。环境科学与技术41:555 - 562。http://dx.doi.org/10.1021/es0610333
Dambacher, J. M., P. C. Rothlisberg和N. R. Loneragan. 2015。定性数学模型,以支持基于生态系统的澳大利亚北部对虾渔业管理。生态应用程序25:278 - 298。http://dx.doi.org/10.1890/13-2030.1
Defeo, O.和J. C. Castilla. 2012。拉丁美洲和加勒比沿海贝业的治理和可治理性:全球化和气候变化的多尺度新兴模式和影响。beplay竞技环境可持续性的最新观点4:344 - 350。http://dx.doi.org/10.1016/j.cosust.2012.05.002
埃斯皮诺萨-特诺里奥,A., M.沃尔夫,I.埃斯佩杰尔和G. Montaño-Moctezuma。2013.利用传统生态学知识改进整体渔业管理:墨西哥南部泻湖生态系统的跨学科建模。生态和社会18(2): 6。http://dx.doi.org/10.5751/es-05369-180206
埃辛顿,t.e., L. Ciannelli, S. S. Heppell, P. S. Levin, T. R. McClanahan, F. Micheli, É。E. Plagányi, I. E. van Putten, 2016。海洋渔业的经验主义和模型:推进一门跨学科科学。生态系统: 1 - 8。http://dx.doi.org/10.1007/s10021-016-0073-0
芬克贝涅e . 2014。墨西哥小规模渔业的生存和可持续性:海洋社会-生态系统恢复力的跨尺度检验.论文。斯坦福大学,美国加利福尼亚州斯坦福。(在线)网址:https://searchworks.stanford.edu/view/10701355
菲什威克,P. A.卢克,1991。定性仿真建模与分析.Springer-Verlag,纽约,纽约,美国。http://dx.doi.org/10.1007/978-1-4613-9072-5
富尔顿,E. A. J. S.林克,i.c.卡普兰,M.萨维娜-罗兰,P.约翰逊,C.安斯沃斯,P.霍恩,R.戈顿,R. J.甘布尔,A. D. M.史密斯和D. C.史密斯。2011。海洋生态系统建模和管理的教训:亚特兰蒂斯的经验。鱼和渔业12(2): 171 - 188。http://dx.doi.org/10.1111/j.1467-2979.2011.00412.x
Halpern, b。,c . Longo d·哈迪k·l·麦克劳德j . f . Samhouri S . k .卡托纳k . Kleisner S e·莱斯特·m·Ranelletti j . O ' leary, a . a·罗森博格c·斯卡伯勒·e·r·塞利格b d最好,d . r . Brumbaugh f·S·查宾,l·b·克劳德·k·l·戴利,S . c·多尼c .精灵m . j . Fogarty S·d·盖恩斯k·雅各布森,l·b·卡勒·h·m·莱斯利·e·Neeley d .保利S Polasky Ris,英国圣马丁岛,g . S .石头,r . Sumaila和d·西。2012。评估全球海洋健康和利益的指数。自然488:615 - 620。http://dx.doi.org/10.1038/nature11397
休斯,T. P., D. R.贝尔伍德,C. Folke, R. S. Steneck, J.威尔逊,2005。支持海洋生态系统恢复力的新范式。生态学与进化趋势“,20(7): 380 - 386。http://dx.doi.org/10.1016/j.tree.2005.03.022
基廷格,J. N, E. M.芬克贝纳,N. C.班,K.布罗德,M. H.卡尔,J. E. Cinner, S. Gelcich, M. L.康威尔,J. Z. Koehn, X. Basurto, R.藤田,M. R.考德威尔,L. B.克劳德。2013。促进小型渔业可持续性的社会生态系统研究的新前沿。环境可持续性的最新观点5:352 - 357。http://dx.doi.org/10.1016/j.cosust.2013.06.008
角,k . 2009。模糊认知映射在半定量场景开发中的潜力,以巴西为例。全球环境变化19:122 - 133。http://dx.doi.org/10.1016/j.gloenvcha.2008.08.003
贾斯特斯,j . 2006。循环分析和定性建模:局限性和优点。生物学和哲学21:647 - 666。http://dx.doi.org/10.1007/s10539-006-9050-x
莱恩,华盛顿,2008年。系统动力学中图解的出现和使用:一个关键的帐户。系统研究与行为科学“,25:3-23。http://dx.doi.org/10.1002/sres.826
莱恩,P.和T. M.柯林斯,1985。海洋浮游生物群落网络的食物网模型:一种实验介观方法。实验海洋生物学与生态学杂志94:41 - 70。http://dx.doi.org/10.1016/0022 - 0981 (85) 90049 - 8
Leslie, h.m., X. Basurto, M. Nenadovic, L. Sievanen, K. C. Cavanaugh, J. J. Cota-Nieto, B. E. Erisman, E. Finkbeiner, G. Hinojosa-Arango, M. Moreno-Báez, S. Nagavarapu, S. M. W. Reddy, Sánchez-Rodríguez, K. Siegel, J. Ulibarria-Valenzuela, A. Hudson Weaver, O. Aburto-Oropeza。2015.运行社会生态系统框架以评估可持续性。美国国家科学院院刊112(19): 5979 - 5984。http://dx.doi.org/10.1073/pnas.1414640112
莱斯利,h·M和k·l·麦克劳德。2007.应对实施海洋生态系统管理的挑战。生态学与环境前沿“,5(10): 540 - 548。http://dx.doi.org/10.1890/060093
列文、M. J.福格蒂、S. A.穆拉斯基和D.弗鲁哈迪。2009。综合生态系统评估:为海洋生态系统管理发展科学基础。公共科学图书馆生物学7 (1): e1000014。http://dx.doi.org/10.1371/journal.pbio.1000014
莱文,r . 1974。讨论论文:部分指定系统的定性分析。纽约科学院年鉴231:123 - 138。http://dx.doi.org/10.1111/j.1749-6632.1974.tb20562.x
R.莱文斯和C. J.普恰,1988。生物群落中的控制因素。Coenoses3(3): 149 - 154。
马洪,R. P.麦康尼,R. N.罗伊,2008。渔业管理是一个复杂的适应系统。海洋政策32:104 - 112。http://dx.doi.org/10.1016/j.marpol.2007.04.011
B. G. Marcot, R. S. Holthausen, M. G. Raphael, M. M. Rowland, M. J. Wisdom. 2001。利用贝叶斯信念网络,从环境影响报告中评估土地管理方案下鱼类和野生动物种群的生存能力。森林生态与经营153:29-42。http://dx.doi.org/10.1016/s0378 - 1127 (01) 00452 - 2
马托内,R. L. G. 2009。温带岩礁物种、人口和群落结构的地理变异.论文。斯坦福大学,美国加利福尼亚州斯坦福。(在线)网址:http://gradworks.umi.com/33/51/3351489.html
mcay, b.j, F. Micheli, G. Ponce-Diáz, G. Murray, G. Shester, S. Ramirez-Sanchez, W. Weisman. 2014。墨西哥太平洋沿岸的合作社、特许权和共同管理。海洋政策44:49-59。http://dx.doi.org/10.1016/j.marpol.2013.08.001
麦克拉纳汉,J. C.卡斯蒂亚,A. T.怀特,O. Defeo. 2009。通过促进复杂的社会生态系统来修复小规模渔业。鱼类生物学与渔业综述19:33-47。http://dx.doi.org/10.1007/s11160-008-9088-8
McGinnis, m.d和E. Ostrom, 2014。社会-生态系统框架:初期变化与持续挑战。生态和社会19(2): 30。http://dx.doi.org/10.5751/es-06387-190230
McLeod, K. L., J. Lubchenco, S. R. Palumbi, A. A. Rosenberg. 2005。基于海洋生态系统管理的科学共识声明.Compass, Corvallis,俄勒冈,美国。(在线)网址:http://www.compassonline.org/science/EBM_CMSP/EBMconsensus
墨尔本-托马斯,J., S.沃瑟斯彭,B.雷蒙德,A.康斯特布尔,2012。定性网络分析中模型不确定度的综合评价。生态专著82:505 - 519。http://dx.doi.org/10.1890/12-0207.1
Micheli, F., G. De Leo, C. Butner, R. G. Martone和G. Shester. 2014b.评估多种渔业累积影响的基于风险的框架。生物保护176:224 - 235。http://dx.doi.org/10.1016/j.biocon.2014.05.031
米开利,F. G. De Leo, G. G. Shester, R. G. Martone, S. E. Lluch-Cota, C. Butner, L. B. Crowder, R. Fujita, S. Gelcich, M. Jain, S. E. Lester, B. McCay, R. Pelc,和A. Sáenz-Arroyo。2014一个.支持改善海产品生产做法和成果的全系统方法。生态学与环境前沿“,12:297 - 305。http://dx.doi.org/10.1890/110257
Micheli, F, A. Saenz-Arroyo, A. Greenley, L. Vazquez, J. A. E. Montes, M. Rossetto, G. A. De Leo. 2012。有证据表明,海洋保护区增强了对气候影响的抵御能力。《公共科学图书馆•综合》7 (7): e40832。http://dx.doi.org/10.1371/journal.pone.0040832
蒙托亚,J. M.伍德沃德,M. C.埃默森,R. V. Solé。2009.间接效应在真实的食物网中传播干扰。生态90:2426 - 2433。http://dx.doi.org/10.1890/08-0657.1
E. Morales-Bojórquez, M. O. Muciño-Diaz, J. A. Vélez-Barajas。2008.鲍鱼渔业衰退的分析(石决明fulgens而且h . corrugata)沿着墨西哥下加利福尼亚半岛的中西部海岸。贝类研究杂志27:865 - 870。http://dx.doi.org/10.2983/0730 - 8000 (2008) 27 [865: AOTDOT] 2.0.CO; 2
Morales-Zárate, m.v, s.e Lluch-Cota, E. Serviere-Zaragoza和S. Guzmán del Próo。2011.模拟一个被开发的岩石海岸生态系统:墨西哥托尔图加斯巴伊亚。生态模型222:1185 - 1191。http://dx.doi.org/10.1016/j.ecolmodel.2010.12.011
纳尔逊,E., G.门多萨,J.雷盖兹,S.波拉斯基,H.塔利斯,D. R.卡梅伦,K. M. A.陈,G. C.戴利,J.戈尔茨坦,P. M.卡里瓦,E.朗斯多夫,R.奈都,T. H.里基茨,M. R.肖。2009。在景观尺度上建模多种生态系统服务、生物多样性保护、商品生产和权衡。生态学与环境前沿“,7:4-11。http://dx.doi.org/10.1890/080023
奥斯特罗姆,e . 2009。分析社会生态系统可持续性的一般框架。科学325(5939): 419 - 422。http://dx.doi.org/10.1126/science.1172133
Özesmi, U.和S. L. Özesmi。2004.基于人的知识的生态模型:多步模糊认知映射方法。生态模型176:43 - 64。http://dx.doi.org/10.1016/j.ecolmodel.2003.10.027
Peckham, s.h, D. Maldonado Diaz, A. Walli, G. Ruiz, L. B. Crowder和W. J. Nichols. 2007。小型渔业的副渔获危及濒危的太平洋红海龟。《公共科学图书馆•综合》2 (10): e1041。http://dx.doi.org/10.1371/journal.pone.0001041
检查Pérez-Ramírez, M, M. A. Almendarez-Hernandez, G. Aviles-Polanco, L. F. Beltran-Morales。2015.消费者接受生态标签鱼:墨西哥案例研究。可持续性7(4): 4625 - 4642。
Pérez-Ramírez, M., B. Phillips, D. luch- belda和S. luch- cota。2012一个.发展中国家实施渔业认证的展望。海洋政策36:297 - 302。http://dx.doi.org/10.1016/j.marpol.2011.06.013
Pérez-Ramírez, M., G. Ponce-Díaz,和S. luch- cota。2012b.MSC认证在赋予墨西哥渔业合作社权力方面的作用:红岩龙虾共同管理渔业的例子。海洋及海岸管理63:24-29。http://dx.doi.org/10.1016/j.ocecoaman.2012.03.009
波里诺,C. A., O. Woodberry, A. Nicholson, K. Korb和B. T. Hart. 2007。用于生态风险评估的贝叶斯网络的参数化和评价。环境建模及软件22:1140 - 1152。http://dx.doi.org/10.1016/j.envsoft.2006.03.006
R. S.波默罗伊,B. M.卡顿和I.哈克斯,2001。影响渔业联合管理成功的条件:来自亚洲的教训。海洋政策25:197 - 208。http://dx.doi.org/10.1016/s0308 - 597 x (01) 00010 - 0
Ponce-Díaz, G., A. Vega-Velázquez, M. Ramade-Villanueva, G. Leon-Carballo,和R. Franco-Santiago。1998.墨西哥下加利福尼亚半岛西海岸鲍鱼渔业的社会经济特征。贝类研究杂志17(3): 853 - 857。
Ponce-Díaz, G., W.韦斯曼,和B.麦凯。2009.墨西哥渔业管理的共同责任和参与:南下加利福尼亚的经验教训。Pesca y Conservacion1:1-9。
普西亚,C. J.和R.莱文斯,1986。复杂系统的定性建模:循环分析和时间平均导论.哈佛大学出版社,美国马萨诸塞州剑桥。http://dx.doi.org/10.4159/harvard.9780674435070
Ramírez Sánchez, S., B. J.麦凯,T. R.约翰逊和W.韦斯曼。2011。外科手术,formación我们的社会组织和组织的持久存在ribereña de la península下加利福尼亚。地区y皇家社会23日(51):71 - 99。
Ramsey, D.和C. Veltman, 2005。预测扰动对生态群落的影响:定性模型能提供什么?动物生态学杂志74:905 - 916。http://dx.doi.org/10.1111/j.1365-2656.2005.00986.x
Reum, J. c.p, P. S. McDonald, B. E. ferris, D. M. Farrell, C. J. Harvey, P. S. Levin. 2015。支持双壳类水产养殖生态系统方法的定性网络模型国际内燃机学会海洋科学杂志72(8): 2278 - 2288。http://dx.doi.org/10.1093/icesjms/fsv119
理查德森,1986年。因果循环图的问题。系统动力学评估2:158 - 170。http://dx.doi.org/10.1002/sdr.4260020207
Schlager, E.和E. Ostrom, 1992。产权制度与自然资源:概念分析。土地经济学68(3): 249 - 262。http://dx.doi.org/10.2307/3146375
谢泼德,s.a, J. R. Turrubiates-Morales和K. Hall, 1998。墨西哥La Natividad鲍鱼渔业的衰落:过度捕捞还是气候变化?beplay竞技贝类研究杂志17:839 - 846。
谢斯特,2008年。小规模渔业的可持续性:利用参与性研究方法分析生态系统影响、捕捞行为和空间管理.论文。斯坦福大学,美国加利福尼亚州斯坦福。(在线)网址:http://gradworks.umi.com/33/32/3332993.html
谢斯特,G. G.和F.米开利,2011。小型渔业的保护挑战:陷阱和刺网的副渔获物和对生境的影响。生物保护144:1673 - 1681。http://dx.doi.org/10.1016/j.biocon.2011.02.023
斯坦eck, R. S., T. P. Hughes, J. E. Cinner, W. N. Adger, S. N. Arnold, F. Berkes, S. A. Boudreau, K. Brown, C. Folke, L. Gunderson, P. Olsson, M. Scheffer, E. Stephenson, B. Walker, J. Wilson,和B. Worm. 2011。缅因州龙虾渔业的高经济价值创造了一个镀金陷阱。保护生物学25:904 - 912。http://dx.doi.org/10.1111/j.1523-1739.2011.01717.x
斯蒂尔,a.c., J. F. Samhouri, S. Gray, R. G. Martone, M. E. Mach, B. S. Halpern, C. V. Kappel, C. Scarborough, P. S. Levin. 2016。将专家的看法融入到食物网的保护和管理中。保护信.http://dx.doi.org/10.1111/conl.12245
范利奥斯,c.d和p.p Groumpos, 1999。制造系统管理者建模的软计算方法。智能与机器人系统学报26:389 - 403。http://dx.doi.org/10.1023/A:1008165916707
泰斯赫斯特,J. L., L. T. H.纽汉,D. Rissik, R. A.莱彻,A. J.杰克曼。2007。用贝叶斯网络方法评估澳大利亚新南威尔士州沿海湖泊的可持续性。环境建模及软件22:1129 - 1139。http://dx.doi.org/10.1016/j.envsoft.2006.03.003
维加,2001年。西班牙西班牙语的翻译结果:Panulirus中断下加利福尼亚半岛的中部和佩斯克里亚地区的.调查通报,Pesquera-La Paz国家调查中心,墨西哥。
惠特利,新墨西哥州,2011。甲壳类动物对海洋酸化的生理和生态响应。海洋生态进展系列430:257 - 271。http://dx.doi.org/10.3354/meps09185
伍尔德里奇,S.和T.完成。2004。学习从过去事件中预测大规模珊瑚白化:使用遥感数据、原位数据和环境代理的贝叶斯方法。珊瑚礁23:96 - 108。http://dx.doi.org/10.1007/s00338-003-0361-y
伍顿,2002。复杂生态系统的间接影响:最近的进展和未来的挑战。海洋研究杂志48:157 - 172。http://dx.doi.org/10.1016/s1385 - 1101 (02) 00149 - 1
