

去pdf这篇文章的版本
Pavlowich, T。,A. R. Kapuscinski, and D. G. Webster. 2019. Navigating social-ecological trade-offs in small-scale fisheries management: an agent-based population model of stoplight parrotfish (Sparisoma viride加勒比海珊瑚礁渔业)。生态和社会24 (3):1。
https://doi.org/10.5751/es - 10799 - 240301
导航生态权衡小规模渔业管理:一个基于主体的人口模型,红绿灯鹦嘴鱼(Sparisoma viride加勒比海珊瑚礁渔业)
文摘
鹦嘴鱼(家庭鹦嘴鱼科)消耗macroalgae必不可少的过程维持珊瑚礁的生态健康。他们已经成为几个加勒比地区渔业目标,这种做法规定食物和收入也将珊瑚礁面临风险。一些国家已经禁止鹦嘴鱼丰收,但这将造成巨大的困难,资源贫乏的渔民在一些地方,由于高比例的鹦嘴鱼物种了。这项研究告诉发展和评估选择实现了最大程度的人口最少的重建困难强加给渔民。渔业模型可以帮助比较管理选项没有实际的例子如何管理鹦嘴鱼的数量。我们建立了一个基于主体的红绿灯鹦嘴鱼的种群模型(Sparisoma viride),一个关键的食草动物和雌蕊先熟的雌雄同体,预测生态和社会各种渔业管理选项的结果。我们参数化模型来表示大量捕捞渔业、评估不同的上下文管理选项是最相关的。我们比较几个管理选项选择通过与利益相关者的讨论和渔业管理的科学文献。我们发现所有选项立即开始增加种群生物量。更多的限制性的选择导致了更大的提高稳态生物量和收入,代表一个长期的双赢。然而,所有管理选项也引起的初始货币价值下降前的捕获最终反弹,因此时间之间的权衡短期和长期的经济结果。虽然没有完美的策略,经理和利益相关者可以使用这些信息来权衡成本和收益的不同在不同的生态环境中可能的管理选项。介绍
珊瑚礁为沿海社区提供多个有价值的生态系统服务,但管理来维持这些生态系统服务呈现复杂的挑战。他们维持渔业,为旅游业提供的机会,并保护海岸线(Moberg和Folke 1999)。钓鱼在珊瑚礁上,然而,改变了社区和鱼的结构,因此,这些生态系统功能(Dulvy et al . 2004年,格雷厄姆et al . 2005年,Madin et al . 2010年)。关键生态动态是食草鱼类和海胆藻类保持距离,让珊瑚生长和繁殖的调停珊瑚和macroalgae之间的竞争(休斯1994年Mumby et al . 2006年)。钓鱼可以删除食草动物丰富和改变群落结构,使珊瑚礁的风险成为长满藻类,失去了生物多样性,它定义了他们的结构和功能,和未能提供相同的福利社会,否则他们会提供。因此,保持足够的食草性水平是可持续礁渔业管理的基石(Nystrom et al . 2008年,麦克拉纳罕et al . 2012年)。
食草鹦嘴鱼是一个重要的社会因素(家庭鹦嘴鱼科)可以弥补大部分渔民的捕捉(2010年Sabetian Pavlowich和Kapuscinski 2017),构成真正的挑战规范鹦嘴鱼收获不会造成不良影响渔民的生计。在加勒比完成禁止鹦嘴鱼捕捉渔业一直主张(Bellwood et al . 2004年,杰克逊等人。2012年,罗杰斯et al . 2015年)最大化的机会让这些生态系统coral-dominated状态(2006年Mumby Bozec et al . 2016年)。然而,一个完整的禁令也将引起足够的经济困难使这种方法在许多加勒比沿海社区的居民社会上站不住脚的其他一些可行的生计选项(威尔逊2011年cin, et al . 2016年)。在多米尼加共和国,我们研究的实证部分,2年禁止鹦嘴鱼是在2017年6月(Ministerio de中部社会y recurso项目一直2017)。手工渔民在多米尼加共和国报道大幅削减总赶鹦嘴鱼禁令实施以来(t . Pavlowich个人观察)。
Bozec et al。(2016)创建了一个人口模型的加勒比鹦嘴鱼和珊瑚礁动态发现的鹦嘴鱼提取了能够维持生态系统崩溃成一个受水藻控制状态。他们的模型主要基于广泛的人口和红绿灯鹦嘴鱼的生活历史数据(Sparisoma virideunfished珊瑚礁的博内尔岛(van罗阿et al . 1996一个,b)。他们检查渔业生态系统动态范围非常广泛,从钓鱼到食草动物种群动态珊瑚吸积率。他们发现,保护珊瑚礁需要低捕捞死亡率鹦嘴鱼,和禁止使用鱼陷阱达到足够的保护这些物种和食草性。他们的工作提供了一个重要的一步了解钓鱼的方式影响珊瑚礁生态系统以及如何设法保持生态弹性。
我们的目标是推进这一研究探索其他的性能在其他生态环境管理选项,因为我们知道资源管理必然会改变在个案基础上(奥斯特罗姆2007)。我们的三个研究目标,在这个目标,如下:
- 描述鹦嘴鱼人口和捕鱼行为大量捕捞加勒比海珊瑚礁。
- 创建一个人口模型功能像一个过度开发的鹦嘴鱼礁在当前渔业条件。
- 预测和比较的短期和长期的生态和经济效益实现许多不同的潜在的渔业管理条例。
达到第一个目标,我们研究了鱼社区和手工捕鱼行为的渔业生态系统在多米尼加共和国。实现第二个目标,我们修改我们之前创建一个模型,这是基于以前公布的鹦嘴鱼模型(Bozec et al . 2016年)。我们我们的研究范围集中在钓鱼和单个鹦嘴鱼物种的种群动态,这样我们可以探索更多的替代监管选项(Weijerman et al . 2015年)。达到最终的目的,我们使用这个模型来预测和比较七替代渔业法规的影响,包括小说的建议只收获末期公鱼。我们设计了两个方面的分析直接监管的影响使本研究适用于小规模渔业在高度依赖渔业社区。首先,我们的渔民的收入作为主要维度包括比较。第二,我们探索如何红绿灯鹦嘴鱼种群和渔民的收入变化实施后给定的监管,从当前形势下,通过直接干扰,直到新的平衡测定。这种方法承认金融对渔民的影响日常生活的重要性,当试图建立可持续的渔业系统。
方法
我们之前开发的基于代理模型用于红绿灯鹦嘴鱼(Sparisoma viride)人口(Pavlowich et al . 2018年)作为我们的研究的基础。模型建立在先前的模型用于探索的影响钓鱼雌雄同体的鱼类种群和红绿灯鹦嘴鱼的动态人口2004(西德尼和甜菜一个,b奥法雷尔et al . 2015年,Bozec et al . 2016年)。我们参数化模型红绿灯鹦嘴鱼大量手工渔业捕捞,使用收集的数据在早上好男人,多米尼加共和国。我们遵循标准的协议来描述基于代理模型,即。、概述、设计概念和细节,整个方法(格林et al . 2010年)。
实验数据收集
我们的当前状态评估当地的鱼类种群在早上好男人鱼使用水下视觉普查技术进行社会调查(麦克拉纳罕et al . 2007年,朗et al . 2010年)。我们进行横断面调查,30米长4米宽在珊瑚礁栖息地的深度(1 - 25米)在整个早上好男人渔业区。横断面位置选择较大的采样区域内随机选择故意实现代表的报道栖息地的多样性和钓鱼的地方。调查是基于深度分层栖息地在3区:0 - 3米,3 - 8 m, 8-25 m。我们进行了66年2013年在珊瑚礁栖息地横断面和47 2014年横断面。总结这些数据时,我们纠正不相称的抽样工作在珊瑚礁栖息地类型相对于每种生境类型占用的面积。我们发现鱼的长度频率在每个栖息地类型,每个频率乘以面积比例的比值给定栖息地类型的栖息地类型的比例每年发生在我们抽样,然后总结所有栖息地类型的长度频率。例如,鱼观察深层礁地区算作2.77 2.40 2013样品和鱼在2014年的样本,而每个鱼在浅水算作2014年0.19和0.21 2013年鱼(表1)。我们聚合鱼5厘米垃圾箱来匹配其他红绿灯鹦嘴鱼人口研究报告的数据(van罗阿et al . 1996一个奥法雷尔et al . 2015)。我们计算和绘制平均长度调整频率为每年每5厘米大小类,然后绘制总体均值(n = 2),平均值较低,较高的平均值(图1,降低面板)。我们计算了性别比例比2014年观察到的初始阶段末期鱼鱼,唯一我们收集的数据对个人的生活阶段。这些数据被调整为前面描述的分层,占调查不足15厘米和所有的鱼被认为是不成熟的,无论是最初的——还是末期不管颜色。
我们也收集数据种类和大小渔民的日常抓住和利用这些数据模型渔民的大小选择性。这包括识别的物种和测量叉长度每一鱼费舍尔在一天。我们每天记录16的实例从2014年的10个不同的spearfishers。唯一的机会收集这些数据在不影响渔民的正常活动是乘船从天降落点的捕鱼区。因此不是随机抽样但基于方便。我们有意寻求从尽可能多的不同的渔民捕捉数据。在早上好男人,spearfishers实践自由潜水和压缩机潜水。我们无法测量捕获从压缩机后勤约束的潜水员,因为伴随这些渔民。先前的研究在捕鱼观光早上好男人表明压缩机潜水员通常访问更深层,有时更大的鱼。然而,两种类型的渔民在相同的市场环境,和压缩机潜水员抓住相同类型的鱼,也许更多的人,因为自由潜水员(Pavlowich和Kapuscinski 2017)。
估计渔民的收入,我们指定的货币价值相同的方式鱼买家在早上好男人。价格每重量支付鱼是由鱼的大小和种类(Pavlowich和Kapuscinski 2017)。同样大小的分类适用于所有物种,但某些物种可以进入价格类别比其他人更有价值。鹦嘴鱼,例如,可以是“第三类”,这是最宝贵的(RD 37美元;2014年多米尼加比索),或“二等”,这是价值超过第三类和不到头等舱(RD 52美元)。个人超过20厘米叉长度被认为是二等,任何小的是第三类。没有官方规模下限,虽然非常小的鱼(∼15厘米)沮丧,有时拒绝买家的早上好男人。我们表达的价格每公斤多米尼加比索。
概述
目的
模型的目的是探索和比较各种渔业法规的影响人口指标和捕获特性的信号灯鹦嘴鱼的上下文中大量捕捞渔业生态系统。分析的焦点是人口从一个过度开发条件重建的大小和短期和长期影响渔民的收入从这个物种。
状态变量和鳞片
模型有一个等级水平:个人的鱼。状态变量:所描述的个人身份号码,叉长、体重、年龄、和生活阶段。生命阶段水平青少年,初始阶段、过渡阶段和晚期阶段。初始阶段鱼被认为是女性,男性末期鱼。
没有空间组件模型,除此之外,它代表1公顷的珊瑚礁。我们创建的上下文中大量捕捞渔业生态系统通过调整模型模块表示生活史的各个方面。
人口特征是个人总共的数量在每个生命阶段,并在不同大小的类;的性别比例(女:男);生物质能的每个人生阶段的所有鱼类和鱼;意味着成熟鱼的长度;和产卵的潜力比(SPR;女生物量/女生物质时F= 0)。抓鱼的数量描述在每个时间步,鱼的重量,和鱼的市场价值。
和调度过程概述
模型运行在每周的时间步骤。在每个时间步内,五个模块按照以下顺序进行处理:增长,生命阶段过渡,成熟,死亡率,和招聘。所有个人,在一个随机的顺序、工艺模块,然后继续下一个模块在一个随机的顺序。操作的顺序是非常重要的过程,而不是为别人。生命阶段过渡和成熟必须发生在增长,因为转换是基于体型。如果经济增长之后生命阶段过渡或成熟,有些鱼会为他们的身体大小不正确的人生阶段。死亡率和招聘可能发生之前或之后增长和生命阶段过渡并产生相同的结果。
设计概念
出现:种群动态,即。,changes in population size and length and life-phase distributions, emerge from the combination of larval recruitment, somatic growth, life-phase transitions, and mortality. Fishing mortality is an especially important factor because it imposes variable mortality rates on fish of different sizes and life phases.
遥感:鱼可以感觉自己的大小和生活阶段,以及所有其他成熟鱼的大小。我们不知道的机制。评估可能会通过视觉线索;物理交互与其他鱼类,如追逐、尖酸刻薄的,等等;或两者(1975年巴洛,曼迪et al . 2006年)。一条鱼的大小和生活阶段相对于其他人群的影响一个人的生命阶段转换和易感性钓鱼。
交互:生命阶段转换是基于个体的大小相对于其他社会群体。确切的机制,确定哪些鱼改变生活阶段,当这个物种不为人知,但它是基于认为涉及大小的社会等级制度和建立和维护由鱼之间的交互。
特性转化:模型代表生命阶段过渡,运动,和死亡概率,因此随机。增长和招聘的过程是确定的。
观察:我们观察到模型结果在种群层面,包括鱼的分布在subpopulation-level组。
细节
初始化
我们开始模拟人口的10例,10个初始阶段的鱼,和10个末期鱼。我们钓鱼参数调到捕鱼的早上好男人(见进一步解释细节:参数估计),允许模型运行直到种群生物量和渔民的收入稳定(3年或156周)。然后我们记录的人口特征,抓住以下2年来(104周)来表示“之前”地位之前申请的一个渔业法规。
子
子的生活历史过程形成这个人口模型的骨架。这些过程的公式是选择最好的可用的信息。对一些人来说,文学提供了一个好的理解的一般过程和红绿灯鹦嘴鱼。对于其他人来说,我们不得不使用简单的配方和估计参数值。描述的参数化程序,子模型描述后,有好的信息在一些流程和人口特征约束的参数值可能导致预期结果,加强我们对估计的信心。
(我)繁殖
模型假定常数幼虫招聘。,没有stock-recruitment函数。珊瑚礁科学家经常做出这种假设在鱼类种群模型(Kellner et al . 2010年,奥法雷尔et al . 2015年),尽管招聘不同暂时(1998年Tolimieri Sponaugle et al . 2006年)和可能至少部分由当地繁殖(舒尔茨1994年考恩,绿色et al . 2015年)。因为科学家尚未发现stock-recruitment函数珊瑚礁鱼类,不断招聘仍是最合适的建模方法繁殖。在这个模型中,一个固定数量的1厘米的青少年(n = 6)进入人口在每一个时间步。
(2)体细胞的生长
个人成长是一个函数的年龄之前,冯Bertalanffy增长方程:
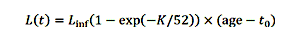 |
(1) |
在哪里l正的最大大小,K是增长系数,t0理论年龄鱼长度为零,虽然长度从来没有真正等于零。增长率除以52把每周的增量。
我们使用以下length-weight关系将鱼的长度,即。在厘米,叉长度,鱼体重,即。,total live weight in kilograms (Froese and Pauly 2017):
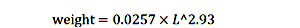 |
(2) |
(3)成熟和性变化
鱼进入模型作为青少年,然后过渡到初始阶段,最后,到终端的阶段。成熟的初始阶段发生时自动鱼成为超过15厘米(van罗阿et al . 1996b)。
的机制,提示初始阶段鱼过渡到终端红绿灯鹦嘴鱼的生活阶段是未知的。然而,加勒比鹦嘴鱼已被证明过渡在人口规模更小的尺寸和结构改变的钓鱼(Choat et al . 2003年,霍金斯和罗伯茨2004)。因此,我们假设个体的概率过渡从最初的终端阶段是一个函数的大小,和过渡在任何给定的概率大小可以改变结构的人口规模变化。我们前面讨论过的结果与一个固定的和三个灵活的规则,可能在红绿灯控制变性鹦嘴鱼(Pavlowich et al . 2018年)。我们没有发现大的差异的不同灵活的规则,所以我们只使用其中的一个。初始阶段的鱼有一个终端阶段过渡到基于概率的成年鱼的人口比例比他们小。变性的概率计算在给定的长度根据西德尼和甜菜(2004年提出的方程b)作为可能的性变化规律:
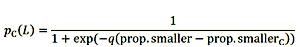 |
(3) |
一个个体的长度(l)确定鱼的相对大小的人口(prop.smaller),还有一些鱼类的比例有50%的机会转变(prop.smallerC)。因此,转变的概率在给定长度变化较大和较小的比鱼的数量长度的变化。我们定义问随着速率的大小获得转变的概率。当鱼被选中过渡到终端阶段,它变成了“过渡”阶段4周,然后成为终端阶段。
(iv)自然死亡率
鱼在模型中经验死亡率从自然原因和钓鱼。自然死亡率有两个组件,捕食鱼很小,衰老当鱼成为老(奥法雷尔et al . 2015)。Bozec et al。(2016)描述了由于自然原因死亡的概率与两个逻辑方程,我们采用模型:
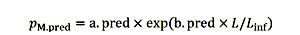 |
(4) |
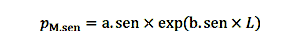 |
(5) |
一个地方。pred尺度捕食死亡率和b。pred决定它速度随大小。为简单起见,我们建模衰老死亡的概率,因为长度的函数,假定年龄和长度有关。一个参数。森和b。森控制程度和速度的死亡率,因为衰老是强加给鱼。
(v)捕捞死亡率
模型使用瞬时捕捞死亡率,F死亡的概率,计算每个鱼因为钓鱼。这个变量封装钓鱼的各个方面的压力,如渔民的数量,每个费舍尔提交多少时间钓鱼,齿轮功效,和鱼的比例遇到追求加特纳(萨拉斯,2004)。我们不要试图解剖这些方面在现实生活中钓鱼,但承认他们的贡献。在模型中,每年瞬时捕捞死亡率除以52把每周的时间步骤。大小和生命阶段选择性修改钓鱼压力来确定死亡的个体的概率。
大小选择性范围从0到1,一个是完全容易捕鱼和死亡的概率等于给定的捕捞压力水平。方程是物流功能:
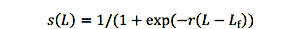 |
(6) |
在哪里lf是鱼的大小有50%的机会被选中,然后呢r是一个速度参数,控制曲线的陡峭。
生命阶段可以选择三个值:任何生命阶段可用(青少年除外),终端,或只初始。如果鱼的生命阶段可以采取的渔业,其死亡的概率如下:
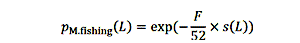 |
(7) |
如果鱼的生命阶段或大小是禁止收获,因为钓鱼死亡的概率是零。注意,在结果中,我们把模式结果与捕捞死亡率作为一个完全的概率选择每年鱼类死亡。这样做可使渔业模型的内部模型方程在一个熟悉的格式,提出了结果在一个更通用的,易于理解的格式。瞬时捕捞死亡率转化为年度钓鱼死亡率按照下列方程(哈登2011):
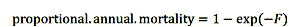 |
(8) |
模型随机选择为每个鱼的死亡率函数应用在每一个时间步,即捕捞死亡率,那么自然死亡率,反之亦然。这避免了问题与假设一个总是先于其他有关。
我们假设渔民没有弥补减少的可用性一定大小和生活阶段通过改变选择性或增加捕鱼工作。不变的假设为非选择性渔具选择性工作得相当好,如吉尔网或鱼陷阱。然而,这种假设可能违反,或多或少,选择性齿轮类型,即捕鱼观光,multispecies渔业,让渔民目标个体鱼的能力。同时,能力目标个体鱼或返回多余的水活鱼,事实上,一个必要元素的最小尺寸和terminal-phase-fish-only法规是有效的。关于补偿通过努力调整,我们没有信息来评估渔民是否会改变他们钓鱼的努力,以应对鹦嘴鱼法规的变化。虽然渔民直接比例更大的努力,不受监管的物种,这样做不会改变我们的结果感兴趣的:红绿灯鹦嘴鱼生物量和他们收获的价值。因此,我们不包括任何机制来增加捕捞死亡率上可用人口当别人成为禁止。
参数估计
我们估计参数值复制红绿灯鹦嘴鱼的大小结构和鱼类密度早上好男人代表一个高度捕捞礁和情节的轨迹恢复从一个现实的起点。参数化模型的第一步是识别大小分布的最重要的特征。其中包括以下定义的属性早上好男人人口:(1)大多数鱼都长10 - 15厘米,(2)分布在大小类越来越多的个人0到15厘米之间,和(3)有相对较少的鱼在25 - 30厘米的类和一些没有鱼超过30厘米。
我们手动搜索模型的参数空间找到参数组合的生产数量与前面描述的特征早上好男人人口因为这三个特征不能被减少到一个单一的指标的拟合优度(格林et al . 2005年)。这涉及到调整四个模型参数以提高适合早上好男人渔业:新兵进入人口的数量在每个时间步(n.recruits),两个参数定义的大小和坡度捕食(死亡率曲线。pred b.pred),捕鱼的数量放在人口压力。前三个参数确定的形状大小分布的小鱼,和钓鱼的压力决定了大个体的分布。我们没有数据来评估死亡率是否因衰老或性变化规律将改变在人口大量捕捞,因此我们从以前的参数化维持这些变量不变(Pavlowich et al . 2018年)。
我们2014年捕捉数据的曲线来描述的大小选择性渔业。为此,我们利用非线性最小二乘符合对数曲线西德尼和甜菜(2004后捕获数据一个)。渔民经常抓鱼是20厘米叉长度(图2)。Twenty-centimeter鱼占15.6%,我们认为任何大小或更大的完全选择(即。、选择= 1.0)。这个假设渔民遇到了其他较小的鱼捕鱼区,但没有抓住这些更小的鱼,因为他们优先目标更大的鱼。我们进一步假设没有大小超过20厘米是大量的捕捉,因为它们更丰富,不是因为渔民没有目标这些尺寸。然后我们满足选择性函数直方图数据如图2所示。最优参数值r= 1.02和长度选择50% = 16.8厘米。
校准招聘、少年死亡,死亡率和钓鱼
我们系统地测试,发现参数组合产生相当好的适合我们观察到的人口,基于先前所描述的主要特征。招聘和控制青少年死亡率的两个参数,一个。pred和b。pred,结合控制鱼的数量在0到15厘米之间。根据死亡率参数的值(一个。pred和b。pred), recruitment could be between 4 and 6 individuals per week and generate reasonable frequencies for the 0- to 5-cm size class. The mortality parameters partially controlled the number of fish in this first size class, as well as frequencies between 5 and 15 cm. We found that the y intercept, a.pred, did best around 0.5 or below. The fit was, of course, dependent on the value of b.pred, which, when paired with a.pred = 0.5 and 6 weekly recruits, performed well when set to −1.75. These were our final parameter values used in the model. With the first part of the size distribution reasonably well fit, we sought to find the level of fishing pressure that would recreate the right-hand side most accurately when paired with the empirically derived size-selectivity parameters and the previously fixed senescence parameters (a.sen and b.sen). Instantaneous annual fishing mortality had to be set to 2.5 to truncate the distribution around 30 cm and have approximately 17 fish in the 25- to 30-cm size class, 2 of our criteria for fitting to the observed size distribution. Together, these parameter values (Table 2, “calibration”) produce a size structure with similar characteristics to those we observed in Buen Hombre. We present a sensitivity analysis on these parameters in the结果。
钓鱼的实验
我们确定了管理选项通过正式和非正式的访谈和与渔民,经理,和一个当地的非政府组织(NGO), AgroFrontera, 2012年和2016年之间(威尔逊et al . 2016年),以及利用这个理念从珊瑚礁管理文学(Heppell et al . 2006年,Bozec et al . 2016年)和我们以前的工作(Pavlowich Kapuscinski 2017 Pavlowich et al . 2018年)。我们确定了四种类型的管理,三我们指定两个级别的强度,导致7个潜在的管理选项。表3给出了干预措施及其细节。
我们测试每个干预通过运行模拟了5年的捕鱼参数估计早上好男人;5年是足够的时间来稳定人口规模和结构。然后我们干预申请另一个5年的模拟。估计早上钓鱼条件的模型稳定后男人第一个3年,之后,我们计算的平均值和标准偏差2年内代表preintervention种群生物量的价值观。我们开始应用管理干预后,我们允许模型3年适应新的捕鱼政权和计算平均2最后几年的模拟。我们重复每个模拟3次因为可变性之间运行很低(见变化模型在图3),从这些模拟,我们追踪种群生物量和渔民的捕捞的价值完整的模拟。这允许我们检查管理选项的影响。我们也计算其他的最终值的结果指标,有助于澄清对人口的影响和捕捉:成年鱼的平均长度;女性的比例生物量当捕捞女性生物量unfished时,即产生潜在的比率(SPR;Nadon et al . 2015); number of fish caught; and weight of fish caught. The value of the catch was highly variable each week, so we used the 6-month moving average of each run to describe its trajectory. We calculated the mean catch value before and after the management intervention the same way as we did for biomass. Additionally, we calculated the time it took for the moving average to return to prior levels after implementing the intervention. We did this because the longer catch value is depressed, the harder it will be on fishers and the less likely fishers will be to comply with a regulation.
敏感性分析
我们测试了模型的灵敏度的关键变量通过改变参数值的基值,然后评估人口和收获成果的关系(Bozec et al . 2016年)。我们随机选择参数值在±25%的基值,绘制他们反对人口规模(图4,上面板)和捕捉价值(下半部分)。我们包括价格变量,因为他们经常变化,和一些利益相关者认为它们是一个点的杠杆改变渔民的行为(f·v·佩顿,AgroFrontera执行主任,个人沟通)。
结果
当前渔业状况在早上好男人
我们观察到在2013年2013年和146年206红绿灯鹦嘴鱼。红绿灯鹦嘴鱼种群的大小结构在早上好男人是定性非常不同于观察unfished博内尔岛(图1)将大量捕捞渔业。人口在早上好男人有更少的大型鱼类,和鱼的频率最小的大小类倾向于增加招聘之后,而不是减少第一15 - 20厘米。我们承认,人口规模和结构以早上好男人抽样的时候可能是暂时的,也就是说。,他们不可能代表一个常数,稳定的状态。我们的观察来自连续2采样季节/ 2年,而数据从博内尔岛(van罗阿et al . 1996一个)是在过去的四年,在此期间人口相当稳定。其他出版红绿灯鹦嘴鱼的大小分布共享的特点,我们的数据或数据从博内尔岛。Koltes特克斯和凯科斯群岛(1993)描述了人口结构类似于在早上的男人:小鱼相对较少,其次是最大频率在中间通过最大尺寸大小和频率递减。Paddack et al。(2009)报道,佛罗里达的大小分布更类似于博内尔岛,有许多新员工,通过中间大小频率下降,另一个峰上大小类。我们观察到性别比例为2.42(初始阶段比终端阶段[IP: TP])在2014年。
渔业干预实验
总人口生物量、人口规模和平均长度最终增加模拟3年后实现的几乎每一个干预(图3、表4)。唯一的例外是,平均长度下降略低于每年3个月封闭的季节。封闭的季节,显示所有干预措施,除了生物量增加后立即实施。生物量增加一段1 - 2年,然后来到一个稳定的状态,周围有一些变化,在每一个新的渔业的监管。季节关闭到位时,生物量迅速上升,然后急剧下降时,渔业再次开放。这种模式持续开启和关闭之间的渔业骑车季节。生物量的大小增加干预之间的差异很大,从一个多增长500%只增加了38%。制定一个最低30厘米的大小限制了最大的鱼类生物量增加preintervention的水平高出518%。捕捞死亡率减少一半,只有收获末期个人也产生大量增加的生物量高于当前的水平,分别为341%和174%。创建3 -或6个月封闭的季节,设定一个最低20厘米的大小,或减少钓鱼仅导致生物质只有小幅上涨10%。其他人口指标的长期反应,人口规模和平均长度,显示类似的模式生物。
最终的平均值的捕捉也增加了每个管理选项(图3),因为重建种群最终提供了更大的鱼,是值得更多的钱(表4)。有很多可变性围绕捕捉价值超过总生物量,导致我们使用6个月移动平均线代表趋势。最大的经济收益来自干预措施也导致增加鱼类生物量最大。至少30厘米的大小捕捉价值增加了177%,和抓只末期鱼翻了一倍的价值。第二大增加捕捉价值,降低捕捞死亡率50%,只有一半的增值抓只末期鱼,和其他干预措施导致更小的增加价值。封闭的季节产生的振动值,为生物一样,导致整体价值的增加平均在整个一年。的场景下30厘米的最小尺寸,减少捕鱼50%,鱼的数量减少,但增加的规模和价值超过弥补了捕获稍少。与总生物量、经济收益没有立即意识到对于任何监管。首次实施干预时捕捉价值开始下降。在所有情况下,捕捉价值最终恢复并超过preintervention水平。
或许最大的挑战涉众希望实现这些法规中可以看到初始捕获价值下降。时间赶上值恢复preintervention水平取决于渔业的严重程度限制。涨幅最大的干预导致生物质和捕捉价值,设定一个最低30厘米的大小,还需要最长的时间赶上水平恢复。捕捉价值低于它最初超过20个月,这包括许多周没有抓住。抓只末期鱼,另一个表现良好的监管,抑制捕捉价值为1年。钓鱼的压力降低了50%保持值为1年4个月。设定一个最低20厘米的大小导致几乎没有初始捕获下降值和相应的小生物量的增加。
SPR增加每个监管下(表4)。渔业科学家经常考虑SPR在0.2和0.3之间过度捕捞的阈值(雷斯特雷波et al . 1998年)。在我们的模拟,实现没有监管,20厘米的最小大小,捕捞死亡率降低10%,或关闭季节3或6个月长导致spr低于0.3。至少30厘米的大小产生最好的SPR的结果,近尽可能多的雌性生物完全unfished人口。
敏感性分析
模型是更敏感的招聘和捕捞死亡率比其他生活史参数(图4)。员工的数量和大小的选择(fork.length.f50)有很强的积极与人口规模的关系,和每年捕捞死亡率有强烈的逆关系。参数,早期死亡率。pred和b。pred,也显示出明确的关系,但是它们对人口规模的影响是很小的。价格没有影响人口规模,因为模型的简化假设没有反馈捕捞压力或fisher行为。这种假设对于渔民的情况下有一些生计选择和几个选项的鱼丰收,一样在早上好男人。这样的约束,费舍尔价格变化将是有限的。捕捉价值对每个变量在同一个方向人口规模,只影响包含更多的变化,就像在我们管理仿真结果。有点令人惊讶的是,价格变量仍然抓住价值没有发挥多大作用。这说明鱼的可用性和大小如何影响渔业收入远远超过每磅的价格。
讨论
我们试图确定管理选项,最大限度地恢复的红绿灯鹦嘴鱼生物量同时最小化负面影响渔民的生计。所有的选项显示这些生态和社会目标之间的权衡。选择特定类型的干预是基于利益相关者和仔细检查优先级目标和可接受的水平的具体结果。这加强了科学家和决策者之间的合作的重要性,在开发一个管理计划的过程。
预测管理的结果
管理选项我们测试现在的渔民和经理双赢和权衡根据时间考虑。我们发现每一个限制我们测试导致股票状态和改善渔业收入从长远来看。然而,我们还发现时间权衡产生最好的结果的干预长期把最难的金融影响渔民在短期内。这些干预措施产生最大的初始蘸捕捉价值和需要最长的时间赶上值恢复到preintervention水平。事实上,长期福利之间的冲突和短期牺牲通常出现在自然资源管理(Mardle Pascoe 2002查宾et al . 2009年)。
各种管理选项的分析可以帮助利益相关者识别和比较具体的选项之间的权衡,确定方法来减轻负面影响不可避免的权衡,从而做出更明智的决定追求一个给定的选项,选项,或修改这些选项。总体而言,该模型预测,法规,限制某些类型的鱼,例如,size restrictions and the terminal-only restriction, produced better economic results for similar levels of population biomass gain than did restrictions on effort. Of particular interest, the terminal-only restriction yielded lower biomass gains than reducing fishing by 50%, but fishing only the terminal phase led to substantially higher catch value. When fishing pressure was reduced by 50%, biomass in all sizes increased to some extent, but this option does not address the current practice of catching small fish that are still undergoing fast growth. Furthermore, reducing fishing pressure by 50% would likely deeply disrupt many fishers’ livelihoods, whether half of fishers were excluded, all fishers were allowed to fish half as much, or half of the fishing area was converted to a no-take zone. In practice, these 3 ways of reducing fishing mortality by 50% would have substantially different outcomes. They would also require tremendous effort to reduce fishing by participation, time, or space. Had we tested other levels of reductions in fishing mortality, we may have found a level that could lead to good population rebuilding with less disruption to income. Our goal, however, was to explore a broad range of regulation types, not to find the optimal level of any one intervention. Doing so better matches the scope and level of detail needed in heavily fished Caribbean reefs where management is currently weak or just beginning.
terminal-phase-only监管经济也表现在重要方面比其他可能的干预措施。相比30厘米的大小限制,它允许渔民继续钓鱼,虽然少了鱼总的来说,虽然人口重建。此外,平均捕获价值回到preintervention水平8个月前比30厘米的限制。只有末期钓到鱼时,释放钓鱼人口死亡率的初始阶段组件允许最大限度增加因为没有损失钓鱼。这反过来又提供了一个更大的鱼可以过渡到终端阶段和增加鱼的频率在中上的大小(即类。,20 - 25厘米,25 - 30厘米的类)。因此,末期的鱼类补充的速度增加,末期鱼更大、更有价值。
封闭的季节比表现得差3和6个月terminal-phase-only限制并与其他管理选项。因为捕捞死亡率仍然很高在那几个月里捕鱼被允许,人口和收入波动强烈。关闭6个赛季大幅提高了平均人口规模,但三个月关闭很难增加人口规模。缺乏积极成果的禁渔期来源于假设,根据经验的观察,这在今年全部或大部分物种产生,没有年度迁移或聚合(Koltes罗伯逊和华纳1978年,1993年,范·罗阿et al . 1996b)。由于这个原因,人口规模迅速下降,均衡的方向现状人口规模在捕捞死亡率,每当钓鱼被允许,即。6到9个月。关于生计,渔民可以适应这个物种的时候不会是可用的,尤其是战略实施不重叠与其他季节因建立龙虾(Panulirus argus)和海螺(Strombus牡蛎)。然而,它也会让另一个挑战的渔业社区生计选择很少。确定社区的偏好将需要进一步咨询。
实施条例以增量的方式可能会创建一个平滑过渡为渔民被迫处理暂时减少了。渔业收入在短期内不会大幅下降,如果经理首先实现terminal-only规则,然后低30厘米的大小限制一段时间后(例如,2年后)。允许末期鱼被抓住会立即降低捕捞死亡率但仍然允许一些收获,而30厘米的最小大小最初将渔民的鱼没有任何的办公处。它还将允许下的人口大幅恢复terminal-phase-only规则,这将使一个30厘米的大小限制破坏性更小的渔民的生计。此外,初期和末期个体之间的差异是渔民更容易区分视觉(Pavlowich et al . 2018年)比鱼是否大于或小于一个特定的大小。这种优势的terminal-only监管将更容易跟随水下spearfishers选择决策和任何监控,如政府官员或其他渔民,在降落点。然而,它不会减少红绿灯鹦嘴鱼渔民的经济依赖。相反,它可能会使人产生强烈的依赖,使它更加难以最终实现全面禁止这个物种。任何新近实施的捕鱼限制可能会破坏一些人支持自己的能力独自钓鱼(cin 2014)。涉及政府和非政府组织加强监管,应伴以支持替代生计和潜在的短期金融安全网方案。 For example, temporary subsidies could help fishers during the model’s predicted 1-year time period, after which population recovery is predicted to restore catch value. Additionally, the multispecies nature of coral reef fisheries provides fishers with opportunities to catch species other than parrotfish to compensate for the decrease in fish availability. However, this would raise concerns about the impacts on other species whose ecological roles might be important for system functioning.
模型估计性别比例约2.5 (IP: TP)在钓鱼的水平压力和大小选择估计早上好男人。这个结果匹配的性别比例2.42,我们观察到在早上好男人。然而,其他红绿灯鹦嘴鱼捕捞的人群有更高的性别比例与unfished人口(van罗阿et al . 1996b霍金斯和罗伯茨2004年,奥法雷尔et al . 2016年)。尽管先前的研究显示,高度捕捞珊瑚礁性别比例失衡,相对正常的性别比例在早上好男人质疑无处不在的这一现象是如何和鹦嘴鱼的程度可以维持一个典型的性别比例,同时进行渔业剥削。显然,我们有限的理解机制推动变性的鹦嘴鱼需要更多的研究才能准确预测的影响这些物种的繁殖行为生态学上钓鱼。
Multispecies上下文
我们调查了一个物种,但是我们发现通常适用于大多数的动力学鹦嘴鱼物种。我们希望大多数物种的数量定性反应类似的类型规定我们测试,虽然影响的大小可能会有所不同。例如,redband鹦嘴鱼(Sparisoma aurofrenatum)可能反应少大小限制,因为它的最大尺寸大小仅略大于鱼类的收获在早上好男人(森林和保利2017)。因此,大多数redband鹦嘴鱼已经能够活到成熟,和一个小比例的人口受到捕捞死亡率的影响。另一个考虑是,物种较慢的增长速度预计将从最初恢复失去的价值更慢,因为这将需要更长的时间来开发更大的鱼。另外,每个鹦嘴鱼物种可能有不同的性变化规律,从而产生不同的反应,尤其是在性别比例。加勒比鹦嘴鱼物种变化性在不同的大小取决于人口的结构,因此,所有物种应该能够在一定程度上补偿增加末期鱼捕捞死亡率(Choat et al . 2003年,霍金斯和罗伯茨2004)。然而,鹦嘴鱼和其他雌蕊先熟的雌雄同体的研究显示了令人担忧的性别比例倾斜在捕捞种群(西德尼和甜菜2004bHeppell等。2006年,奥法雷尔et al . 2016年)。我们的模拟表明,这可能是因为捕捞死亡率高的初始阶段鱼除了强烈的影响钓鱼在终端阶段。密切监测应该伴随任何实现life-phase-based监管的钓鱼,避免生殖失败如果性改变在这些物种不能跟上的速度提取(Heppell et al . 2006年,西德尼et al . 2008年)。
另一个管理考虑应用简单的规定我们的影响被认为是所有物种在multispecies珊瑚礁渔业。每个规则建模应该降低捕捞死亡率的物种应用,从而帮助重建利用种群的生物量。然而,随之而来的捕捉价值下降后立即每个监管的应用程序(见图3)将有一个更大的负金融对渔民的影响如果它包含来自多个物种的收入损失。应用30厘米的大小限制等规定,所有物种,都但是关闭整个渔业在早上好男人在一段时间内,使这一战略社会不能存活的。然而,将限制鹦嘴鱼就可以导致渔民切换目标物种,因此,寻求其他的鱼以更大的强度(cin 2011)。虽然最近更多关注于珊瑚礁健康食草性的必要性(Bellwood et al . 2004年,休斯et al . 2010年),其他官能团无疑扮演了一个重要的角色在维持生态系统的结构和弹性。因此,这将是重要的监测渔业和鱼社区响应以避免意想不到的后果。超越新法规的影响渔民的生计本身,渔民捕鱼可能转变压力的程度选择,nonherbivorous物种提供了一个重要的原因,探索金融弥合策略可能会限制渔民的临时抓收入的下降。
未来的工作
几个领域的研究可以拓展我们的研究范围和提高其预测的准确性和精度。整合其他鹦嘴鱼物种,然后non-parrotfish物种到相同的建模框架有助于阐明社区而不是人口的影响提出管理干预措施。这种扩张将会需要更复杂的子的物种相互作用,社区动态,渔民的捕鱼行为,特别是关于选择决策和能力改变他们的努力时,禁止某些类型的鱼。这些新的子的复杂性也需要经验观察,如我们之前工作spearfishers的觅食行为(Pavlowich和Kapuscinski 2017),让他们可信。同样,一种改进的理解性的线索和机制变化鹦嘴鱼可以验证当前的模型或提出改进建议。此外,长期观察鹦嘴鱼种群早上好男人会进一步支持或提供机会改进的模型参数,提高模型预测的准确性。或许最重要的是,实施的任何测试法规和监控生态、经济、和行为反应系统将提供关键数据模型验证和改进。
结论
我们使用了一个基于主体的建模平台,汇集最有效的信息在生态和经济上重要的红绿灯鹦嘴鱼和预测潜在的管理干预措施的积极和消极的结果。我们定制的模型和问题的生态环境管理最需要:小规模礁渔业在发展中国家。我们发现限制捕捞只末期鱼可以提供过渡性战略对这个物种从一个完全缺乏监管更严格的和生态恢复的限制。高资源依赖的情况下,退化生态系统,和低水平的治理、利益相关者和科学家应该首先寻求策略使生态和社会情况更好,不一定最优,步骤复杂的路径向可持续渔业管理。
确认
我们要感谢社区的早上好男人和房地美博士佩顿的非政府组织AgroFrontera同时开展这一研究的支持和友谊。我们感谢中国环境和自然资源,“五分镍币消费品展,“让我们为开放工作在该地区和讨论关于当地的渔业管理。我们感谢莫莉威尔逊,吉尔布里顿,莎拉令他们的援助。我们欠了谢奥法雷尔和朱尔斯·范·罗阿提供数据从他们的研究在博内尔岛红绿灯鹦嘴鱼。
文献引用
西德尼,s . H。,T. Ish, M. Key, A. D. Maccall, and M. Mangel. 2008. The importance of incorporating protogynous sex change into stock assessments.《海洋科学83 (1):163 - 179。
西德尼,s . H。,和M. Mangel. 2004一个。size-selective渔业的影响和精子的股票动态限制在变性鱼。渔业公告102 (1):1-13。
西德尼,s . H。,和M. Mangel. 2004b。变性规则,股票动态,和雌蕊先熟的股市spawning-per-recruit性能的措施。渔业公告103 (2):229 - 245。
巴洛,g . w . 1975。社会生物学的四个波多黎各鹦哥鱼类(鹦嘴鱼科)。海洋生物学33:281 - 293。https://doi.org/10.1007/BF00390566
Bellwood, d R。,T. P. Hughes, C. Folke, and M. Nyström. 2004. Confronting the coral reef crisis.自然429:827 - 833。https://doi.org/10.1038/nature02691
Bozec Y.-M。,S. O’Farrell, J. H. Bruggemann, B. E. Luckhurst, and P. J. Mumby. 2016. Tradeoffs between fisheries harvest and the resilience of coral reefs.美国国家科学院院刊》上的美利坚合众国113 (16):4536 - 4541。https://doi.org/10.1073/pnas.1601529113
查宾,我。f·S。,G. P. Kofinas, and C. Folke. 2009.生态系统管理的原则:resilience-based自然资源管理在一个变化的世界。施普林格,纽约,纽约,美国。
Choat, j . H。,D. R. Robertson, J. L. Ackerman, and J. M. Posada. 2003. An age-based demographic analysis of the Caribbean stoplight parrotfishSparisoma viride。《海洋生态发展系列246:265 - 277。https://doi.org/10.3354/meps246265
cin, j . e . 2011。在礁渔业生态陷阱。全球环境变化21 (3):835 - 839。https://doi.org/10.1016/j.gloenvcha.2011.04.012
cin, j . 2014。珊瑚礁的生计。当前的舆论环境的可持续性7:65 - 71。https://doi.org/10.1016/j.cosust.2013.11.025
Dulvy: K。,R. P. Freckleton, and N. V. C. Polunin. 2004. Coral reef cascades and the indirect effects of predator removal by exploitation.生态学通讯7 (5):410 - 416。https://doi.org/10.1111/j.1461-0248.2004.00593.x
森林,R。,和D. Pauly. 2017.FishBase。定量游泳Inc .,拉古纳、菲律宾。(在线)网址:https://www.fishbase.org
格雷厄姆:a . J。,N. K. Dulvy, S. Jennings, and N. V. C. Polunin. 2005. Size-spectra as indicators of the effects of fishing on coral reef fish assemblages.珊瑚礁24 (1):118 - 124。https://doi.org/10.1007/s00338 - 004 - 0466 - y
绿色,a . L。,A. P. Maypa, G. R. Almany, K. L. Rhodes, R. Weeks, R. A. Abesamis, M. G. Gleason, P. J. Mumby, and A. T. White. 2015. Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design.生物评价90:1215 - 1247。https://doi.org/10.1111/brv.12155
格林,V。,U. Berger, D. L. DeAngelis, J. G. Polhill, J. Giske, and S. F. Railsback. 2010. The ODD protocol: a review and first update.生态模型221 (23):2760 - 2768。https://doi.org/10.1016/j.ecolmodel.2010.08.019
格林,V。,E. Revilla, U. Berger, F. Jeltsch, W. M. Mooij, S. F. Railsback, H.-H. Thulke, J. Weiner, T. Wiegand, and D. L. DeAngelis. 2005. Pattern-oriented modeling of agent-based complex systems: lessons from ecology.科学310 (5750):987 - 991。https://doi.org/10.1126/science.1116681
哈登,m . 2011。在渔业中建模和定量方法。第二版。查普曼和大厅,佛罗里达州博卡拉顿,美国。
霍金斯,j . P。,和C。M. Roberts. 2004. Effects of fishing on sex-changing Caribbean parrotfishes.生物保护115 (2):213 - 226。https://doi.org/10.1016/s0006 - 3207 (03) 00119 - 8
Heppell, S。,S. A. Heppell, F. C. Coleman, and C. C. Koenig. 2006. Models to compare management options for a protogynous fish.生态应用程序16 (1):238 - 249。https://doi.org/10.1890/04 - 1113
休斯,t, p . 1994。灾难、相移和大规模退化的加勒比海珊瑚礁。科学265 (5178):1547 - 1551。https://doi.org/10.1126/science.265.5178.1547
休斯,t . P。,N. A. J. Graham, J. B. C. Jackson, P. J. Mumby, and R. S. Steneck. 2010. Rising to the challenge of sustaining coral reef resilience.生态学与进化的趋势25 (11):633 - 642。https://doi.org/10.1016/j.tree.2010.07.011
杰克逊,j . b . C。,M. Donovan, K. Cramer, and V. Lam. 2012.加勒比海珊瑚礁的状况和趋势:1970 - 2012。腺,瑞士。
柯尔尼,j·B。,S. Y. Litvin, A. Hastings, F. Micheli, and P. J. Mumby. 2010. Disentangling trophic interactions inside a Caribbean marine reserve.生态应用程序20 (7):1979 - 1992。https://doi.org/10.1890/09 - 1217.1
Koltes, k . h . 1993。方面的红绿灯鹦嘴鱼的生殖生物学和社会结构,Sparisoma viride在格兰特,特克斯和凯科斯群岛,B.W.I.《海洋科学52 (2):792 - 805。
朗,j . C。,K。W. Marks, P. A. Kramer, P. R. Kramer, and R. N. Ginsburg. 2010.AGRRA协议版本5.4。海洋研究和教育基金会,大松键,佛罗里达,美国。
Madin, e . m . P。,S. D. Gaines, J. S. Madin, and R. R. Warner. 2010. Fishing indirectly structures macroalgal assemblages by altering herbivore behavior.美国博物学家176 (6):785 - 801。https://doi.org/10.1086/657039
Mardle, S。,和S. Pascoe. 2002. Modelling the effects of trade-offs between long and short-term objectives in fisheries management.环境管理杂志》65 (1):49 - 62。https://doi.org/10.1006/jema.2001.0518
麦克拉纳罕,t·R。,S. D. Donner, J. A. Maynard, M. A. MacNeil, N. A. J. Graham, J. Maina, A. C. Baker, J. B. Alemu I, M. Beger, S. J. Campbell, E. S. Darling, C. M. Eakin, S. F. Heron, S. D. Jupiter, C. J. Lundquist, E. McLeod, P. J. Mumby, M. J. Paddack, E. R. Selig, and R. van Woesik. 2012. Prioritizing key resilience indicators to support coral reef management in a changing climate.《公共科学图书馆•综合》7 (8):e42884。https://doi.org/10.1371/journal.pone.0042884
麦克拉纳罕,t·R。,N. A. J. Graham, J. Maina, P. Chabanet, J. H. Bruggemann, and N. V. C. Polunin. 2007. Influence of instantaneous variation on estimates of coral reef fish populations and communities.《海洋生态发展系列340:221 - 234。https://doi.org/10.3354/meps340221
Ministerio de中部社会y recurso项目一直》2017。五分镍币环境apuesta关于proteccion de especies码头。Mediante la Resolucion 023/2017。Ministerio de中部社会y recurso项目一直,圣多明各多米尼加共和国。
支持F。,和C。Folke. 1999. Ecological goods and services of coral reef ecosystems.生态经济学29 (2):215 - 233。https://doi.org/10.1016/s0921 - 8009 (99) 00009 - 9
Mumby, p . j . 2006。利用食草动物的影响(鹦嘴鱼科)的动态加勒比海珊瑚礁。生态应用程序16 (2):747 - 769。https://doi.org/10.1890/1051 - 0761 (2006) 016 (0747: TIOEGS) 2.0.CO; 2
Mumby, p . J。,J. D. Hedley, K. Zychaluk, A. R. Harborne, and P. G. Blackwell. 2006. Revisiting the catastrophic die-off of the urchinDiadema antillarum在加勒比海的珊瑚礁:从仿真模型弹性方面的新见解。生态模型196 (2):131 - 148。https://doi.org/10.1016/j.ecolmodel.2005.11.035
芒,p . L。,P. M. Buston, and R. R. Warner. 2006. Diversity and flexibility of sex-change strategies in animals.生态学与进化的趋势21 (2):89 - 95。https://doi.org/10.1016/j.tree.2005.10.020
Nadon, m . O。,J. S. Ault, I. D. Williams, S. G. Smith, and G. T. DiNardo. 2015. Length-based assessment of coral reef fish populations in the main and northwestern Hawaiian Islands.《公共科学图书馆•综合》10 (8):e0133960。https://doi.org/10.1371/journal.pone.0133960
Nystrom, M。,N. A. J. Graham, J. Lokrantz, and A. V. Norström. 2008. Capturing the cornerstones of coral reef resilience: linking theory to practice.珊瑚礁27 (4):795 - 809。https://doi.org/10.1007/s00338 - 008 - 0426 - z
奥法雷尔,S。,B. E. Luckhurst, S. J. Box, and P. J. Mumby. 2016. Parrotfish sex ratios recover rapidly in Bermuda following a fishing ban.珊瑚礁35 (2):421 - 425。https://doi.org/10.1007/s00338 - 015 - 1389 - 5
奥法雷尔,S。,R. Salguero-Gómez, J. M. van Rooij, and P. J. Mumby. 2015. Disentangling trait-based mortality in species with decoupled size and age.动物生态学杂志84 (5):1446 - 1456。https://doi.org/10.1111/1365 - 2656.12399
奥斯特罗姆,e . 2007。超越灵丹妙药的诊断方法。美国国家科学院院刊》上的美利坚合众国104 (39):15181 - 15187。https://doi.org/10.1073/pnas.0702288104
Paddack, m . J。,S. Sponaugle, and R. K. Cowen. 2009. Small-scale demographic variation in the stoplight parrotfishSparisoma viride。鱼的生物学》杂志上75 (10):2509 - 2526。https://doi.org/10.1111/j.1095-8649.2009.02451.x
Pavlowich, T。,和A. R. Kapuscinski. 2017. Understanding spearfishing in a coral reef fishery: fishers’ opportunities, constraints, and decision-making.《公共科学图书馆•综合》12 (7):e0181617。https://doi.org/10.1371/journal.pone.0181617
Pavlowich, T。,d·g·韦伯斯特,和A. R. Kapuscinski. 2018. Leveraging sex change in parrotfish to managed fished populations.人类世的Elementa:科学6 (1):63。https://doi.org/10.1525/elementa.318
雷斯特雷波,v R。,G. G. Thompson, P. M. Mace, W. L. Gabriel, L. L. Low, A. D. MacCall, R. D. Methot, J. E. Powers, B. L. Taylor, P. R. Wade, and J. F. Witzig. 1998.技术指导在实现国家标准的预防性的使用方法1的Magnuson-Stevens渔业保护和管理行为。NOAA技术备忘录NMFS-F / SPO-31。美国商务部,国家海洋和大气管理局,国家海洋渔业服务,美国马里兰州银泉。
Robertson d R。,和R. R. Warner. 1978.性模式labroid鱼类的加勒比海西部II:鹦哥鱼类(鹦嘴鱼科)。史密森学会贡献动物学255。史密森学会出版社,华盛顿特区,美国。https://doi.org/10.5479/si.00810282.255
罗杰斯。,A. R. Harborne, C. J. Brown, Y.-M. Bozec, C. Castro, I. Chollett, K. Hock, C. A. Knowland, A. Marshell, J. C. Ortiz, T. Razak, G. Roff, J. Samper-Villarreal, M. I. Saunders, N. H. Wolff, and P. J. Mumby. 2015. Anticipative management for coral reef ecosystem services in the 21st century.全球变化生物学21 (2):504 - 514。https://doi.org/10.1111/gcb.12725
Sabetian, a . 2010。鹦嘴鱼渔业和种群动态:一个案例研究从所罗门群岛。论文。詹姆斯库克大学,澳大利亚昆士兰州汤斯维尔。
加特纳萨拉斯,s和d》2004。渔民:管理的行为动力学的影响。鱼和渔业5 (2):153 - 167。https://doi.org/10.1111/j.1467-2979.2004.00146.x
舒尔茨,e . T。,和R. K. Cowen. 1994. Recruitment of coral-reef fishes to Bermuda: local retention or long-distance transport?《海洋生态发展系列109:15-28。
Sponaugle, S。,K。Grorud-Colvert, and D. Pinkard. 2006. Temperature-mediated variation in early life history traits and recruitment success of the coral reef fishThalassoma bifasciatum在佛罗里达群岛。《海洋生态发展系列308:1-15。
Tolimieri: 1998。小环境特征之间的关系,招聘和成人丰富红绿灯鹦嘴鱼,Sparisoma viride在三个空间尺度上。《海洋科学62 (1):253 - 268。
范·罗阿,j . M。,J. P. Kok, and J. J. Videler. 1996一个。当地人口结构和密度变化的雌蕊先熟的食草动物Sparisoma viride。环境生物学的鱼47:65 - 80。https://doi.org/10.1007/BF00002380
范·罗阿,j . M。,F。J. Kroon, and J. J. Videler. 1996b。社会和食草鱼类的交配系统Sparisoma viride:一男与博士组。环境生物学的鱼47:353 - 378。https://doi.org/10.1007/BF00005050
Weijerman, M。,E. A. Fulton, A. B. G. Janssen, J. J. Kuiper, R. Leemans, B. J. Robson, I. A. van de Leemput, and W. M. Mooij. 2015. How models can support ecosystem-based management of coral reefs.海洋学的进展138 b: 559 - 570。https://doi.org/10.1016/j.pocean.2014.12.017
威尔逊,M。,T. Pavlowich, and M. Cox. 2016. Studying common-pool resources over time: a longitudinal case study of the Buen Hombre fishery in the Dominican Republic.中记录45 (2):215 - 229。https://doi.org/10.1007/s13280 - 015 - 0688 - y
