

去pdf本文版本
茵斯-戈尔德,a.a., T. Pavlowich, M. Heinichen, M. C. McManus, J. McNamee, J. Collie和A. T. Humphries, 2021年。利用食物网和人类行为耦合模型探索渔业中的社会-生态权衡。生态和社会26(2): 40。
https://doi.org/10.5751/ES-12451-260240
利用食物网和人类行为耦合模型探索渔业中的社会-生态权衡
摘要
海洋渔业是一个受复杂生态过程和人类相互作用驱动的社会-生态系统。以生态系统为基础的渔业管理需要了解生物和社会两方面的组成部分,如果其中任何一项被排除在外,就可能发生管理失败。尽管两者都很重要,但由于很难将人类行为纳入模拟模型,大多数研究都专注于描述生物不确定性,而不是更好地理解人类行为的影响。在本研究中,我们以纳拉甘塞特湾(美国罗德岛州)的渔业为案例研究,展示如何使用耦合模型来表示社会-生态系统中食物网和渔民之间的相互作用。纳拉甘西特湾是一个商业性的饲料鱼渔场,例如大西洋鲱鱼(推喇奴Brevoortia)及供捕食者(如条纹鲈鱼(马龙saxatilis)及蓝鱼(Pomatomus saltatrix).为了探索这两种渔业之间的权衡,我们创建了一个食物网模型,然后将其与休闲渔民的行为模型相结合,创建了生态系统的动态社会-生态表征。在独立的食物网模型和耦合的社会-生态模型中,对到2030年的鱼类生物量进行了预测,结果强调了在建模中结合渔民的行为会如何导致生态系统的变化。我们考察了模型输出是如何随三个属性而变化的:(1)饲料鱼的商业捕捞情景,(2)休闲渔业中掠食性(鱼食性)鱼的丰度-捕获量的关系,以及(3)休闲渔民失去信心的比率(称为“满意度损失”)。饲料鱼类的商业捕捞量越高,鱼食性鱼类的生物量越低,但对休闲捕捞鱼食性鱼类的数量或休闲渔民的满意度影响最小。丰度-渔获量关系和满足损失率对鱼类生物量、休闲渔获量和休闲渔人满意度均有显著影响。目前,缺乏特定于空间和位置的fisher行为数据限制了我们模型的预测性使用。然而,我们的建模框架表明,通过使用基于代理的建模,渔人行为可以成功地纳入一个耦合的社会-生态模型,我们的结果强调,它的纳入可以影响生态系统动态。由于渔民的决策和生态系统可以相互影响,因此社会对生态系统变化的反应应该明确地纳入生态系统模型,以改善基于生态系统的渔业管理工作。介绍
自然资源管理者和政策制定者面临着如何在一个耦合的社会-生态系统中维持资源的使用(Schlüter et al. 2012, Guerrero et al. 2018)。这些相互关联的系统通过结合自然成分和社会、经济、政治和文化因素,提供了各种生态系统服务(Boyd和Banzhaf, 2007)。管理这些社会-生态系统是困难的,因为它们具有适应性、异质性、多尺度和时变的特性(Liu et al. 2007)。海洋渔业是一项重要的全球产业,代表了社会-生态系统的许多特性,其中包括复杂的环境和生态过程,以及巨大的人类影响(Fulton et al. 2011)。实现渔业可持续性需要了解生物因素,即鱼类种群,以及社会因素,包括沿海社区生计和经济利益(Guerrero等,2018年)。
渔业科学努力将对生态系统的物理、生物和生态因素的理解纳入其中,包括使用捕捉物种间动态的食物网模型。Ecopath with Ecosim (EwE)是一个食物网模型,用来模拟生物的能量流和生物量(Polovina 1984, Christensen and Pauly 1992)。EwE的一个指导原则是能量质量平衡,这意味着对于模型中的每一个群体,能量的去除,即捕食或捕鱼,必须与消耗的能量相平衡(Coll et al. 2009)。组间能量平衡用两个线性方程表示,分别对应生产和消耗。Ecosim使用时间序列数据,并将这些线性方程重新表示为时变微分方程,从而使Ecopath的静态、能量平衡的食物网快照在时间上是动态的。通过Ecosim,用户可以模拟食物网如何应对捕鱼死亡率或初级生产等因素的变化(Coll等人2009年,Heymans等人2016年)。EwE被广泛用于评估渔业对生态系统的影响(Pauly et al. 2000)。例如,Buchheister et al.(2017)使用EwE模拟不同的大西洋鲱鱼(推喇奴Brevoortia),并发现它们的天敌大西洋条纹鲈鱼(马龙saxatilis)对这些变化最为敏感。事实上,当鲱鱼以最大可持续产量捕捞时,条纹鲈鱼的产量和生物量预计将下降约60% (Buchheister et al. 2017)。虽然EwE可以模拟捕鱼努力的变化,但它没有办法包括来自动态捕鱼者行为的反馈。
当渔业仅根据单一物种的需求进行管理,而忽视了社会背景时,管理失败就经常发生(Cooper and Jarre 2017)。特别是,考虑到资源使用者的行为经常与管理者和经济理论的预期不同,人类行为已被确定为渔业科学和基于生态系统的渔业管理的一大不确定性来源(Fulton et al. 2011)。在一个耦合的社会-生态系统中,这种不确定性可以通过系统之间的联系而复合,创造一个不可靠的模型(Cenek和Franklin 2017)。尽管需要纳入社会和经济目标,大量研究仍继续集中于增加生态知识。对渔业管理中人类行为的解释已导致其被纳入最近的一些建模工作(Ono等人2017年,Matsumura等人2019年,Kaemingk等人2020年)。基于agent的建模(ABM)是一种很有前途的人类行为表征方法,非常适合基于生态系统的渔业管理(Burgess等,2020年)。在ABMs中,一套规则管理着自主的“代理”(例如,个人或渔船)如何与彼此以及被建模的环境进行交互(Macal和North 2005)。abm在资源开采系统中尤其有用,比如渔业,在这些系统中,agent和环境本身都存在异质性(Nolan et al. 2009)。尽管这是一种有用的工具,但在渔业科学和经济学中使用ABMs的情况并未得到充分利用(Cooper和Jarre 2017)。
本研究的总体目标是建立一个社会-生态耦合模型,以了解渔人动态因素对生态结果的影响程度。具体来说,我们研究了我们的模型预测如何对三个可能影响社会-生态相互作用的因素作出反应:(1)商业渔业收成的变化,(2)休闲渔业捕捞可能性的变化,以及(3)休闲渔民失去信心并改变其行为的速度。响应变量包括鱼的生物量、休闲渔业捕获的鱼的数量,以及休闲渔民在一个季节结束时的满意度。为此,我们创建了一个基于个体的休闲渔民和鱼类动态模型(以下称为“渔民行为模型”),并将其与EwE模型(以下称为“食物网模型”;据我们所知,这是将这些类型的模型联系起来的第一次尝试,尽管每种模型都已经建立了研究工具。我们在不同的场景下运行耦合的EwE-ABM模型(以下称为“社会-生态模型”),并描述模型运行的差异。在这项工作中,我们预测生物生物量在不同的情况下,以证明我们的模型的使用。然而,目前缺乏fisher行为参数的定量数据,因此我们测试了一个假设值的范围。因此,我们的目标不是预测纳拉甘西特湾鱼类生物量将如何随时间变化。相反,我们展示了一个框架来连接两种已建立的建模方法,并展示了包括人类决策的动态如何改变模型预测,从而影响管理。
方法
研究地点及背景
纳拉甘西特湾(美国罗德岛州)有着悠久的捕鱼和数据收集历史。纳拉干西特湾的第一个定量渔业数据是在19世纪后期收集的,因为陷阱和鱼钩渔民之间的冲突,他们声称鱼类资源正在下降(Oviatt et al. 2003)。这些早期记录显示,捕获的鱼类以溯河洄游物种(鲱鱼[Alosa pseudoharengus,美式shad [Alosa sapidissima,彩虹香鼻[Osmerus mordax])、北方物种如冬比目鱼(Pseudopleuronectes也),以及大西洋鲱鱼等迁徙物种。随着环境条件和物种群落组成的变化,这些以前的优势物种在20世纪后期逐渐减少(Oviatt et al. 2003),其中包括温水物种(Stenotomus chrysops)及蝶鱼(Peprilus triacanthus)变得普遍(Collie et al. 2008)。如今,捕鱼业支撑着罗德岛经济的很大一部分。2016年,罗德岛商业海鲜行业的经济总产值为4.19亿美元,支持了4000多个就业岗位(美世和斯普劳尔2018年)。休闲渔业也发挥着很大的经济作用,2016年销售额为4.12亿美元,收入为1.76亿美元,并支持了4000多个就业岗位(国家海洋渔业局2018年)。在纳拉甘西特湾上游,包括渔民在内的游船用户占所有用户的近三分之二(Dalton et al. 2010)。
纳拉甘西特湾的渔业为探索耦合社会-生态系统中的渔业权衡提供了一个案例研究。以大西洋鲱鱼等饲料鱼为目标的商业渔业主要在格林尼治湾、布里斯托尔港和希望山湾的海湾中上游地区进行(图2)。除了作为商业捕鱼目标外,饲料鱼在纳拉甘塞特湾食物网中发挥着重要作用;它们是下层营养层浮游生物和上层捕食者(如鱼食性鱼类、海鸟和海洋哺乳动物)之间的能量管道(Innes-Gold等,2020年)。由于这种联系,饲料鱼作为目标捕食者物种的主要食物来源,对纳拉甘西特湾休闲渔业非常重要。这些捕食者,包括条纹鲈鱼和蓝鳍鱼(Pomatomus saltatrix),是具有经济和社会价值的热门休闲捕鱼目标(数据查询来自国家海洋渔业局渔业统计司)。尽管由于补充动态,饲料鱼的数量变化很大,但过度捕捞会加剧这些波动,导致过度捕捞的种群对生态系统产生广泛影响,尤其是对饲料鱼捕食者(Essington等,2015)。相反,通过捕鱼的捕食者的开发被证明是级联和影响饲料鱼种群动态(Engelhard et al. 2014)。
饲料鱼作为支持定向商业渔业的角色和为休闲的鱼食性鱼类提供食物的角色之间的权衡给管理带来了复杂的挑战,这不是纳拉甘塞特湾独有的。在整个美国东海岸(Buchheister et al. 2017)和全球范围(Houle et al. 2013),关于捕猎猎物及其捕食者如何相互影响,以及在此背景下如何管理整体捕猎的问题已经被详细研究和讨论(Kuffner 2017, Chase 2019)。尽管这一问题已经在更广泛的地理范围内得到了探索,但纳拉甘塞特湾作为许多鱼类的河口苗圃有着明显不同的动态(孟et al. 2002, Mateo et al. 2011)。此外,过去的这些研究并没有明确地考虑到鱼类的行为如何随着食物网的变化而变化。目前,纳拉甘西特湾的渔业管理人员面临的问题是,在没有适当工具的情况下,饲料鱼的捕捞是如何直接影响该物种的种群动态及其支持的休闲渔业的。
食物网模型
纳拉甘西特湾食物网模型是利用EwE创建的(Innes-Gold等人,2020年)。中上层营养功能类群为鱼食性鱼类、饲料鱼类、底栖动物、肉食性底栖动物、鱿鱼、悬食性底栖动物、养殖贝类和海鸟。较低营养级官能团为沉积食性底栖动物、胶状浮游动物、浮游动物、浮游植物、底栖藻类和碎屑。商业和休闲渔业也派代表参加了会议。在这项研究中,我们主要对饲料鱼(大西洋鲱鱼、鲱鱼、大西洋鲱鱼[Clupea harengus],蓝背鲱鱼[Alosa -],湾凤尾鱼[Anchoa mitchilli]、大西洋月鱼[月之女神setapinnis])及鱼食性鱼类(条纹鲈鱼、蓝纹鱼、夏比目鱼[Paralichthys dentatus),一种食用鱼Cynoscion regalis],多刺角鲨[Squalus acanthias])官能团。食物网模型的官能团生物量数据主要由罗德岛大学海洋学研究生院或罗德岛环境管理部门海底拖网调查提供。产量/生物量(P/B)值为Z,即总死亡率、自然死亡率和捕捞死亡率之和。消耗/生物量(Q/B)值主要来自鱼基(Froese和Pauly, 2019年)和托马斯-布雷的无脊椎动物消耗方程(http://www.thomas-brey.de/science/virtualhandbook/navlog/index.html).生态营养效率(EE)无法在田间测量,因此Ecopath使用所有群体的线性方程来求解(Christensen et al. 2005)。休闲渔业登陆来自美国国家海洋和大气管理局(NOAA)的国家海洋渔业服务查询工具(https://www.st.nmfs.noaa.gov/st1/recreational/queries/)和商业着陆要么是根据NOAA船只旅行报告的数据请求估计的,要么是根据罗德岛州着陆的比例(https://www.accsp.org/).饮食资料来源于多种文献和资料收集方法。食物网模型的强迫函数为浮游植物生物量(g/m²)、养殖贝类生物量(g/m²)和渔业死亡率(F;根据Christensen等人(2005年)和Heymans等人(2016年)的建议,估算了渔业目标功能群的渔获量/生物量。有关模型输入和数据源的更多细节,请参见Innes-Gold等人(2020)。
费雪的行为模型
我们利用NetLogo (Wilensky 1999)在纳拉甘塞特湾的食鱼休闲渔业基础上创建了渔民行为ABM。虽然我们将这个ABM称为“渔民行为模型”,但它包括休闲渔民和鱼类种群动态,详细内容如下。我们的渔人行为ABM使用了概述、设计概念和细节协议(Grimm et al. 2006, 2010, 2020)。
目的和模式
该模型的目标是创建一个纳拉甘西特湾休闲渔业的动态表示,并将其与食物网模型联系起来,并检查渔民行为成分对模型预测的影响。我们将使用这种耦合的社会-生态模型来量化饲料鱼的商业收获、丰度-捕获量关系和渔民满意度损失率如何影响鱼的生物量、休闲渔民捕获的鱼食性鱼的数量及其满意度。
实体、状态变量和规模
第一种类型的代理是鱼。为了反映鱼类种群的动态变化,赋予鱼类agent若干属性。属性包括年龄,长度,生殖状态(所有的开始都是“准备产卵”,产卵后改变为“不准备产卵”,在一年的剩余时间),和性别(假设1:1的性别比例;Terceiro 2010, Sharov et al. 2013)。数据来源于条纹鲈鱼、蓝纹鱼和夏比目鱼,这是三种最常见的休闲捕鱼目标,属于鱼食性鱼类的功能组。必须包括这些种群特征,以便鱼类种群能够恢复和维持自身,而不是随着捕鱼而不断下降。由于这些数值不易从EwE模型中获得,因此均来源于文献。在渔民行为模型中,休闲渔业有两种类型或“品种”:滨岸渔民和渔船渔民。渔民的满意度从2到10不等(Shafer 2007),这决定了他们参与渔业的程度。这个模型代表了纳拉甘塞特湾的整体,没有进一步的空间成分。
过程概述
每个时间步为一周。1-26周是休闲钓鱼的“开放季节”,代表5月至10月,27-52周是“封闭季节”,代表一年中的其他时间。这是假设每年有一半的时间用于休闲钓鱼。在每个每周的时间步中,都有一些鱼剂因自然死亡而先死亡。鱼也会繁殖、生长和衰老。岸上和船上的渔民根据自己的满意度来决定是否去捕鱼。然后,活跃的渔民试图抓住一条鱼。如果他们成功捕到一条鱼,他们决定是把鱼养着还是放着。如果钓到一条鱼,渔夫的满意度就会增加;如果一个活跃的渔夫什么也没钓到,他们的满意度就会下降,这就使得他们在接下来的一周不太可能再活跃起来捕鱼。然后该模型进入下一个每周时间步骤。
设计概念
突出性:鱼类种群动态在模型运行过程中持续,每条鱼的年龄、生长、繁殖和死亡都是由于自然死亡率和捕捞死亡率在每个周时间步。这维持了总体人口结构。
适应:渔夫根据之前时间步骤的捕鱼成功来适应。如果成功了,渔夫就更有可能在接下来的时间步再次捕鱼。
随机性:我们已经用随机概率表示了以下过程:鱼的自然死亡率、释放死亡率、渔民捕鱼的决定、捕获的可能性和保留一条鱼的可能性。
观察:我们观察捕获和饲养的鱼的数量,以及在一个季节结束时种群中剩余的数量。我们还注意到每个季节结束时渔民的平均满意度。
初始化和输入数据
通过将食物网模型预测的2019年食鱼鱼类生物量(在商业捕捞情景中存在差异)转换为鱼类数量(表1)来计算初始鱼类种群大小。为此,将g/m²生物量乘以海湾的面积(380平方公里;2017年纳拉甘西特湾入海口项目)。然后,总生物量被划分为代表不同鱼类大小类别的不同部分(表A1.1,根据NJ鱼类与野生动物部门2010年的条纹鲈鱼数据估算,Sharov等人2013年)。每个大小类别的鱼的平均长度被转换成一个重量使用一个公布的条纹鲈鱼的长度-重量方程(Kimmerer et al. 2005)。每个大小类别中鱼类的总生物量除以该大小类别中个体的重量,就得到了该大小类别中在海湾中鱼类的大致数量。将这些大小类计数总数相加,得到鱼的总数量,并通过在NetLogo中运行时将数值缩小1000倍进行转换。纳入尺寸/年龄结构种群使鱼类具有特定尺寸的增长率和特定年龄的死亡率,这对以尺寸为主导的休闲渔业特别重要。
每条鱼的长度(英寸)是根据泊松分布随机抽取的。40%的鱼种群来自平均5英寸的泊松分布,30%来自平均10英寸的泊松分布,30%来自平均25英寸的泊松分布,这与公布的长度分布近似(NJ鱼类和野生动物部门,2010年,Sharov等人,2013年)。鱼的年龄属性是通过使用Von Bertalanffy方程(方程1)从它们的指定长度计算出来的,其中k是一个生长系数,代表L无穷大的速率,或渐近最大长度(以下为L正),是接近(Froese和Pauly 2019)。对于我们的鱼剂,k设为0.2和L正50英寸。
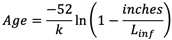 |
(1) |
在所有的试验中,每个渔民的满意度都是通过随机抽取一个均值为5的正态分布分配的(SD = 1),假设渔民的满意度水平存在一定的变化,并且大多数渔民一开始的满意度是中等水平(Arlinghaus 2006, Pascoe et al. 2015)。每一种渔民的初始参与者总数是基于2018年罗德岛每年休闲钓鱼旅行的次数,转换为每周参与,假设所有钓鱼都发生在一年中的6个月(数据查询来自国家海洋渔业服务,渔业统计部门)。得到的结果是,每周有35000次上岸捕鱼和9000次乘船捕鱼,次数分别减少了1000次至35次和9次。每次模型运行都使用相同数量的鱼和渔夫。
子
- 自然死亡率:存在与年龄相关的自然死亡率。鱼的年龄越大,自然死亡的概率就越低。使用的周自然死亡率估计数为0.041(0岁)、0.022(1岁)、0.004(2岁)、0.003(3岁)、0.001(4岁及以上)。鱼的最长寿命为21年。
- 体的生长和成熟:鱼按照Von Bertalanffy生长方程(方程2)生长正设置为50英寸,k设置为0.2。每个时间步鱼龄+ 1周。
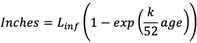 |
(2) |
- 繁殖:5岁以上的雌鱼每个季节可以产卵一次(NOAA渔业2020a, b, c)。考虑到一定的密度依赖性,如果鱼的种群数量小于5000,一条雌鱼产卵12个新成员,如果种群数量为5000 - 15000,则产卵7个新成员,如果种群数量为15000 >,则产卵3个新成员。
- 钓鱼活动:如果在1到10之间产生的随机数小于钓鱼者的满意度,那么他们就会变得“活跃”,并会去钓鱼。因为满意度的最小值是2,所以钓鱼者总是有机会变得活跃,但满意度越低,钓鱼的可能性就越小。
- 决定保留或释放一条鱼:如果渔夫捕获了一条饲养员大小的鱼,称为饲养员,(28-35英寸,目前条纹鲈鱼法规在RI;罗德岛DEM 2020),渔民保留鱼的概率为0.85,释放鱼的概率为0.15。如果他们抓到一条不合法的鱼,他们就把它放了。
- 放生死亡率:放生的饲养员和亚合法鱼类在放生时死亡的几率分别为7%和10%(根据物种特异性研究范围:迪奥达蒂和理查兹1996年、露西和霍尔顿1998年、马尔霍夫和露西1998年、纳尔逊1998年、大西洋州海洋渔业委员会2015年)。
- 丰度-渔获量的关系:渔夫捕获一条鱼的可能性取决于鱼的丰度。在这项研究中,我们使用三种不同类型的渔获丰度关系的渔人行为模型:(1)幂函数,(2)线性关系,(3)静态百分比(图3)。从岸上或陆地捕获一条鱼的概率之间的关系首先被定义为幂函数(公式3,来自Gaertner和Dreyfus-Leon, 2004)。我们创建了四个独立的幂函数,分别表示渔民从船上捕到次合法鱼、从岸上捕到次合法鱼、从船上捕到次合法鱼和从岸上捕到次合法鱼的概率(表A1.2)。
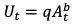 |
(3) |
在这个幂函数(式3)中,Ut为t时刻单位努力捕获量(CPUE),问catchability,一个t充裕在时间t,和b是形状参数。b在输入其他参数后(使用的值可以在表A1.2中找到)。为Ut,我们使用海洋康乐资讯计划的数据进行康乐钓鱼活动,选择“罗德岛内陆”作为我们感兴趣的地区(https://www.st.nmfs.noaa.gov/st1/recreational/MRIP_Survey_Data/).岸上或船上捕获一条或多条鱼食性鱼类(条纹鲈鱼、蓝鲶、夏比目鱼)的次数除以一年的总次数,计算成功捕获一个养鱼人的次数的百分比。将放生一条或多条鱼食性鱼类的海岸或船只航行次数除以一年的总航行次数,计算成功捕获一条亚合法鱼类的航行次数的百分比,假设所有放生的鱼都是亚合法大小的。对于饲养员和从船上捕获的次合法鱼类,使用了2017年的数据。对于饲养者和从岸上捕获的次合法鱼类,近年来的成功率非常低,这就形成了一个在非常低的比率上稳定下来的函数,因此使用了过去25年的平均成功率。对于饲养员而言,可捕性(问)为0.04,离海岸0.02。sublegal鱼,问船上的值是0.08,岸上的值是0.04。这些数值是根据对小口鱼(Micropterus dolomieu)及大口黑鲈(Micropterus salmoides),是鱼钩和鱼线休闲渔业的最佳可用数据(Hangsleben et al. 2013, Wildenhain 2016, Hansen 2018)。时间t (一个t)是通过将2017年的鱼食性鱼类生物量(取自食物网模型时间序列)转换为合法和次合法鱼类数量来计算的,使用上述方法来计算鱼类的起始数量。然后我们解决了形状参数。我们选择用幂函数进行试验,因为它们经常被用来理解观察到的渔获量与物种丰度之间的关系(Hilborn 1985, Gaertner and Dreyfus-Leon 2004)。
线性关系也被用来关联丰度和渔获量(Paloheimo and Dickie 1964, Campbell 2004)。建立了四种线性函数的变化,用于从渔船或岸上方法捕捉亚合法鱼类和饲养员(表A1.2)。捕鱼者和亚合法鱼的线性丰度-渔获量关系是通过定义起点和鱼丰度高点之间的斜率来创建的,在这个点上幂曲线开始趋于平稳,接近一个最大值点,这意味着渔获量不再增加(5000个饲养者,10000条亚合法鱼)。最后,根据海洋康乐资讯计划的数据(表A1.2),以过去25年成功航行的平均百分比来确定丰度与渔获量的静态百分比。采用幂函数部分描述的方法,一次跳闸成功。
- 满意度损失率:如果一次钓鱼不成功,钓鱼者变得沮丧的比率,即零渔获,我们称之为“满意度损失”,直接决定了钓鱼者继续钓鱼的可能性。我们运行了三个不同版本的fisher行为模型来测试模型结果对满意度损失率的敏感性。三种满意度的损失分别是:每周一个积极的渔夫什么也没钓到,损失1分、一半和0分。在所有试验中,如果抓到一条次合法的鱼,满意度会提高0.1分,如果抓到饲养员大小的鱼,满意度会提高1分,因为研究表明,休闲钓鱼者在抓到更多鱼时满意度会提高(Pitman等人,2019年)。
模型校准
我们设置了几个模型参数,使种群在没有捕鱼的情况下保持稳定。当有条件时,我们从以前对条纹鲈鱼、蓝鱼和/或夏季比目鱼的研究中的参数值开始,然后进行增量调整,直到种群既不降为零,也不增至无穷大,并尽可能与观测到的大小分布相匹配(NJ鱼类与野生动物部门2010,Sharov等人2013)。在我们的模型中,每个产卵鱼的新成员数量在3到12之间,取决于鱼的丰度。这一产卵鱼范围的计算方法是将条纹鲈鱼招募的数量除以产卵鱼的数量(Sharov等人2013年),使用2000克的平均产卵鱼重量将产卵鱼公吨转化为产卵鱼数量,这是在已公布的产卵鱼重量范围内(Morse 1981, Kimmerer等人2005,Robillard等人2008)。Martino和Houde(2012)估计条纹鲈鱼的零年龄死亡率为0.068,我们将其降低到0.041,以重现观察到的大约1岁的鱼的数量。通过将条纹鲈鱼瞬时死亡率(大西洋国家海洋渔业委员会2015年)转换为年度死亡率(Gulland 1969年),然后使用A1.2方程(Krebs 2014年)转换为周死亡率,计算其他年龄特定死亡率。1、2、3、4、5和6岁以上儿童的初始周死亡率分别为0.022、0.013、0.009、0.005、0.004和0.003(大西洋国家海洋渔业委员会,2015年)。为了保持长期稳定的种群数量,它们的数量被降低了。其他调整包括将寿命从18.6岁(条纹鲈鱼、蓝鳍鱼和夏季比目鱼的平均寿命;Froese and Pauly 2019年)到21岁,将L.inf提高了40英寸(三个物种的平均水平; Froese and Pauly 2019) to 50 inches, and increasing the K of 0.18 (the average of the three species; Froese and Pauly 2019) to 0.2.
虽然我们是基于未加权平均值建立模型的,但这三个物种(条纹鲈鱼、蓝鱼、夏比目鱼)的调整值在范围内,而且事实上,这三个物种在纳拉甘塞特湾的种群数量并不相等,因此改变初始的未加权估计并非不合理。我们首先将重点放在特定年龄的死亡率数据上,因为这些数据只来自条纹鲈鱼,而蓝鱼和夏季比目鱼的数据是不可用的。为了不把死亡率降低到不现实的水平,我们对其他参数做了轻微的调整,以达到人口稳定。有了这些调整后的值,模型在没有捕鱼的情况下运行了200年,并确定模型是稳定的,表现为种群或大小分布没有增加或减少的趋势。
模型耦合
在这里,我们描述了食物网和渔人行为模型的双向联系,我们将其统称为“社会-生态模型”(图1)。我们的工作始于Innes-Gold等人(2020)描述的EwE食物网模型,从1994年持续到2018年。在单独的试验中,将四种商业收获F值分别添加到2019年作为食物网模型(1994-2018年)的强迫函数的现有时间序列中。所有其他强迫函数保持在2018年的值不变。然后,食物网模型又运行了一年,预测出2020年的鱼食性鱼类生物量。然后,根据条纹鲈鱼的长度分布(表A1.1),将食鱼鱼的总生物量(g/m²)转换为鱼的数量,开始渔人行为模型(表1)。表1显示了每种商业捕捞情景如何导致不同的鱼食性鱼类生物量,因此在渔民行为模型中,鱼食性鱼类的起始数量不同。渔人行为模型运行了一个季节(26周),清除的数量(保留的鱼+死亡的丢弃物)转换为鱼食性鱼的捕捞死亡率(F)。清除的鱼的数量转换为捕获生物量(g/m²),使用与描述的方法相反的方法将g/m²生物量转换为鱼的数量。捕获的生物量(g/m²)然后除以被用作该时间步的渔民行为模型输入的生物量,以计算F值。由于食物网模型中使用的原始食鱼鱼F值是商业和休闲F值的总和,因此根据渔人行为模型计算的新F值被加入到商业F值中,假设从2018年开始预测的商业F值为常数(~0.03)。然后,将合并后的F值返回到强迫函数时间序列中,作为2020年的鱼食性鱼F(所有其他强迫函数保持不变)。 The food web model was then run again for an additional year, through 2021, where the resulting piscivorous fish biomass was used as the input to the fisher behavior model and the process was repeated. This iterative process was applied each year through 2030.
试验类型
商业收获试验
商业收获试验仅在食物网模型中进行。我们模拟了四个场景,以探索不同数量的商业饲料鱼类收获如何影响鱼食性鱼类和其他功能群。饲料鱼捕捞死亡率是食物网模型中使用的强迫函数时间序列,并应用于饲料鱼功能群。我们对饲草鱼捕捞死亡情景的范围进行了测试:零渔获的封闭渔业(F = 0)、现状(食物网模型时间序列中2014-2018年F值的平均值,F = 0.202)、中间情景(现状和极端情景的平均值,F = 1.68)和极端情景(食物网模型时间序列中F值最高的年份为2000年,F = 3.167)。在单独的试验中,从2019年开始输入每个F值,并在未来10年保持不变(例如:2019 - 2029年F = 0)。在每一个食物网模型预测中,其他强迫函数(F表示其他官能基团、浮游植物生物量、养殖贝类生物量)在未来10年保持在2018年的值不变。本试验研究的响应变量为中上层官能团的生物量。
Abundance-catch关系试验
在耦合的社会-生态模型中进行了丰度-渔获量关系试验。这个模型的一个不确定性来源是鱼的数量和渔民成功捕到一条鱼的概率之间的关系,因为这种关系在纳拉甘西特湾还没有被定义。上述三种类型的函数被用于不同的渔民行为模型试验,以预测给定的一次钓鱼活动是否能够成功捕获亚合法或饲养员大小的鱼(图3)。所有这些试验都使用了0.5的满意度损失率。每一项试验(动力试验、线性试验、静态试验)都在4个商业饲料鱼收获场景(共12次试验)中进行,只与同一场景中的其他试验进行比较。每一个具体的试验都在耦合的社会-生态模型中进行了10年(直到2030年)。测试这些不同的丰度-渔获量关系,可以评估对鱼食性鱼类生物量、渔获数量和渔人季终满意度的响应变量对假设关系的敏感性。
满意度损失率试验
满意度损失率试验也在耦合的社会-生态模型中进行。休闲渔民的满意度损失率也是该模型的不确定性来源。这种不确定性可能是必然的,因为满意率直接决定今后的渔业参与和努力。我们进行了三次试验,让渔民在一次不成功的旅行后失去1个满意点、一半满意点或0个满意点,也就是说,没有鱼被捕获。在这些试验中,丰度-捕获关系总是使用幂函数来定义,因为那是零模型。3个试验(1个、一半、0个)分别在4个商业饲料鱼收获场景(共12个试验)中进行,只与同一场景中的其他试验进行比较。每项试验都在耦合的社会-生态模型中进行了10年(直到2030年)。在本试验中,三个响应变量也是鱼食性鱼类的生物量,捕获的鱼的数量和渔人的季末满意度。
分析
在商业收获情景试验中,我们报告了由食物网模型预测的2030年生物生物量的差异。在丰度-渔获量关系试验中,我们详细描述了三个响应变量的线性、静态和动力试验之间的差异:鱼食性鱼类生物量、捕获和饲养的鱼的数量,即“饲养员”和季节结束满意度。对于满意度损失试验,使用相同的三个响应变量的1、一半和0试验之间的差异被描述。未进行统计检验,以避免仿真建模得出不恰当的结论(White et al. 2014)。
结果
商业收获情景试验
在独立食物网模型预测中,所有鱼类类群的生物量在禁止捕捞饲草鱼时最高(图4)。在2030年,极端、中等和现状捕捞情景下的饲草鱼生物量分别为零捕捞条件下的44.9%、64.1%和94.7%(表A1.3)。鱼食性鱼类与它们的猎物(饲料鱼)表现出类似的模式,尽管规模较小(图4)。极端、中等和现状捕捞情景下,鱼食性鱼类的生物量分别为零捕捞时预测生物量的75.4%、81.7%和96.8%(表A1.3)。其他中上营养层组对这些不同的饲料鱼类收获情景表现出一系列的反应;肉食性底栖动物的生物量基本不受影响,而底栖鱼类的生物量随饲料鱼产量的增加而略有下降。大乌贼和小乌贼也以相同的方式响应,饲料鱼产量的增加导致乌贼生物量的降低(图4)。在中等和极端捕捞情景下,饲料鱼和两组乌贼生物量的初始下降幅度最大(图4)。对于所有组,零捕捞情景和现状捕捞情景的预测生物量相似,因为现状捕捞死亡率非常低。
Abundance-catch关系试验
改变渔人行为模型中丰度-渔获量关系的性质会导致所有三个响应变量的差异(图5a)。我们使用幂函数来定义丰度-渔获量关系的模型试验预测了更高的渔获量、更高的渔民满意度和更低的鱼类生物量(图5a, A1.1)。具有线性或静态关系的试验预测捕获的鱼更少,渔民的满意度更低,而随后的生物量更高(图5a)。在某些情况下,即零商业收获,线性和静态试验在响应变量上没有产生很大的差异,而在其他情况下,即中间商业收获,它们产生了很大的差异(图5a)。幂函数试验一般产生的鱼的生物量和捕获的一些鱼与静态或线性试验结果有较大的差异,这些结果往往是相似的。对于赛季末的满意度,这种区别不太明显(图5a)。不同的商业饲料鱼类捕捞情景导致了鱼食性鱼类生物量的巨大差异,较高的捕捞情景导致较低的预测生物量,尽管零捕捞情景和现状捕捞情景的预测生物量相似(图6a)。值得注意的是,收获情景在饲养员捕获的数量和渔民的季节结束满意度方面没有显著差异(图6a)。
满意度损失率试验
满意度损失率对捕获鱼类数量和渔人季末满意度产生了明显不同的结果,但对鱼食性鱼类生物量的区分模式较少(图5b)。满意度损失率为0时,捕获鱼数较高,渔人满意度较高,预计鱼生物量较低;满意度损失率为1时,捕获鱼数较低,渔人满意度较低,预计鱼食性鱼生物量较高(图5b, A1.2)。与之前一样,商业饲料鱼类的产量越高,鱼食性鱼类的生物量越低(图6b)。在收获情景中,饲养员的捕获量和渔人的季末满意度几乎没有显著差异(图6b)。
讨论
通过将食物网模型与渔人行为模型联系起来,我们展示了一种社会生态学方法,该方法可以用于测试社会对环境和人为干扰的反应。我们已经表明,包括人的维度成分可能导致生态变量(即鱼类生物量)的模型预测值存在很大差异。当我们将我们的耦合模型预测与独立食物网模型预测进行比较时,可以看到这一点,独立食物网模型没有包括渔人行为模型。通过将这两个模型耦合在一起,预测的生物量值与仅由食物网模型预测的值不同,无论运行哪种试验类型。这有力地支持了之前关于将人类行为纳入生态系统建模和基于生态系统的渔业管理的呼吁(Hall-Arber等人2009年,Hornborg等人2019年)。之前的EwE模型采用了价值链延伸(Christensen等人2011年)或模拟EwE中的渔业管理策略(Martell等人2002年,Chagaris等人2015年),但据我们所知,EwE没有与ABM或动态渔民行为模型联系起来。在EwE模型和fisher行为的时间动态模型(即ABM)之间建立双向耦合,可以更全面地理解社会-生态系统,并让用户阐明导致系统不同响应的重要机制。这样的框架提供了利用模型探索社会-生态场景的可能性,不仅包括调查人类对生态系统的影响,还包括资源使用者如何对生态系统的变化作出反应。因为管理者规范人类的行为,旨在告知管理的研究和建模工作必须考虑社会领域作为生态系统的一部分。
商业收获的场景
食物网模型表明,目前(现状)低水平的捕捞压力对纳拉甘塞特湾的饲料或鱼食性鱼类生物量没有产生显著的影响。如果有更高的捕鱼压力,即在极端情况下,该模型预计所有三种鱼类和鱿鱼的数量都会减少,但这对休闲渔业捕捞的鱼的数量影响相当小。根据目前的捕获量限制,未来商业捕鱼对食浮游鱼类的压力不太可能达到测试的极限水平。在纳拉甘西特湾模型中,两种大小的鱿鱼在不同的饲料鱼收获情景中都出现了相当大的减少,因为饲料鱼在鱿鱼饲料中占相当大的比例(小鱿鱼27%,大鱿鱼50%)(Innes-Gold等人,2020年)。值得注意的是,与三种鱼类相比,这种食物网模型对两组鱿鱼的精确度都不高,这可能导致预测结果不太可靠(Innes-Gold et al. 2020)。在食物网模型中,当捕食者的首选猎物来源减少时,捕食者会切换猎物,这在一定程度上缓冲了饲料鱼丰收对食物网的影响。允许切换猎物在生态学上是合理的,因为河口捕食者已经多次被证明会根据环境变化中不同的猎物可获得性来改变它们的饮食(Pihl等人1992年,Nobriga和Feyrer 2008年,Szczepanski 2013年;m . Heinichen未出版的手稿).总的来说,饲料鱼的收获情况似乎并不是休闲捕捞的鱼食性鱼的数量或休闲渔民的满意度的主要驱动力。
虽然这些结果表明,目前纳拉甘西特湾的饲料渔业压力没有产生较大的社会或生态影响,但这可能不能推广到其他地方的饲料渔业。研究发现,饲料鱼的收获会对更大规模的捕食者生物量产生影响,具体取决于所关注的物种(Buchheister et al. 2017)。有几个原因可以解释为什么我们对捕食者生物量的影响规模较小。通过在我们的饲料和鱼食性鱼类功能组中分组多个物种,我们无法确定哪个物种有最强的反应。Buchheister等人(2017)发现,条纹鲈鱼对鲱鱼捕捞量的变化反应强烈,而其他捕食者的反应则不那么强烈。由于该EwE模型是针对广泛的生态系统层面的问题设计的,并将条纹鲈鱼与其他食鱼鱼类(如蓝鱼和夏季比目鱼)分组在一起,因此不能用于隔离特定物种的变化。此外,纳拉甘西特湾并不是美国东海岸商业捕鱼最密集的河口。例如,切萨皮克湾有更高的饲料鱼收获水平,并有严格的管理系统,旨在考虑鲱鱼的生态系统服务(SEDAR 2015)。
还需要注意的是,在我们进行的预测中,其他强迫函数(浮游植物生物量、养殖贝类生物量、其他鱼类种群的捕捞死亡率)保持在2018年的水平不变。这是为了隔离饲料鱼收获的影响,并具体探索饲料鱼捕捞压力如何影响生态系统。考虑到浮游植物在EwE模型中作为驱动因素的重要性(Innes-Gold等人,2020年),以及纳拉干西特湾营养物质输入的历史(Nixon等人,2008年),未来的模拟可以探索不同捕鱼压力和浮游植物(作为营养物质输入的代表)的联合影响。例如,初级生产力的减少可能导致饲料鱼的减少,因为浮游生物和饲料鱼之间有很强的联系(Cury等,2000)。考虑到养分减少与初级生产者之间的关系是非线性的(Oviatt等人,2017),这是一个值得未来研究的领域。
Abundance-catch关系
在耦合模型模拟中,改变丰度-渔获量的关系会导致渔获量的变化,进而导致鱼的生物量和渔民满意度的变化。我们之所以选择测试丰度-渔获量关系,是因为在其他休闲渔业中,鱼的丰度越高,渔获量也就越高(Pitman等人,2019年),然而,对于估算这一关系的最佳方法并没有达成一致意见。对于某些物种,每单位努力的丰度和捕获量(CPUE)可能有正比关系,但对于其他单个或分组的物种,CPUE被发现是一个较差的丰度指数(Richards and Schnute 1986, Haggarty and King 2006)。历史上,通常用线性方法来联系丰度和渔获量来评估鱼类(Cooke和Beddington, 1984)。虽然线性方法被质疑为过度简化了这种关系(Peterman和Steer 1981, Bannerot和Austin 1983),但也发现在某些情况下,它的表现比幂曲线更好(Tsuboi和Endou 2008)。幂函数是另一种常见的方法,被认为是线性函数更合适的替代方法(Hilborn 1985, Harley et al. 2001, Gaertner and Dreyfus-Leon 2004)。我们选择加入一个额外的静态关系,因为数量和捕捞之间可能没有关系(Tsuboi and Endou 2008)。对定义的丰度-渔获量关系缺乏共识可能是由于渔具对物种脆弱性的不同(Tsuboi和Endou, 2008)。其他因素也会影响CPUE,如鱼类成熟度、鱼类密度、扫掠面积、年份、季节、渔具和渔民行为(Richards and Schnute 1986, Large 1992)。
我们的工作表明,当存在不确定性时,测试多重丰度-渔获量关系的重要性。在我们的模型中,三种不同的丰度-渔获量关系导致了非常不同的模型结果,因为它们的潜在功能形式。当使用幂函数时,预测的渔获数有时是在线性或静态试验中预测的数的两倍。这种差异导致了渔民在渔季结束时的满意度差异,进而影响了他们继续捕鱼的可能性。相比之下,饲料鱼的商业捕捞情景对鱼食性鱼的捕获数量或休闲渔民的满意度影响较小。这些结果表明,丰度-渔获量关系需要明确界定,以保证模型预测的可信度。因此,我们强调了未来研究的另一个领域:量化纳拉甘西特湾休闲渔民的丰度与渔获量的关系。这种关系的性质因地点和物种的不同而不同(Gaertner和Dreyfus-Leon 2004, Tsuboi和Endou 2008),因此很难将源自一个渔场的关系转移到另一个渔场。影响这种关系的因素既有社会因素,即渔具和行为,也有生态因素,即鱼类数量、密度和种群动态。解释影响渔获率的行为变化尤其具有挑战性,这肯定了未来研究的需要,以减少纯生态环境之外的不确定性(Fulton et al. 2011)。
满意度损失率
在耦合模型仿真中,满意度损失率导致实测响应变量存在较大差异。我们的满意度变量代表了许多影响渔夫决策过程的因素,这些因素决定了他们是否会在给定的时间内捕鱼,捕鱼后满意度会增加(Pitman等,2019年)。在我们的模型中,当渔民很快失去信心,去捕鱼的频率降低时,就会导致捕捞的鱼变少,鱼类生物量增加。另一方面,如果上一次的失败并没有让渔民感到气馁,预计捕获量会翻倍。与丰度-渔获量关系相似,满足损失率导致所有响应变量的变化,而商业捕捞情景主要影响鱼类生物量。由于这种影响,为了有可靠的模型预测,量化渔民退出渔业的速度也很重要。
需要填补的关键数据空白包括发现之前的一次旅行在多大程度上影响渔民的捕鱼决定,以及一次成功的旅行增加的满意度是否大于一次不成功的旅行降低的满意度,这是由一个珊瑚礁休闲渔业的ABM提出的(Shafer 2007)。理想情况下,这些数据应该从纳拉甘西特湾休闲渔业社区的调查中收集,因为休闲渔民的不同行为可能会导致不同地点的数据不同(Mackay等,2020年)。一些有用的调查问题包括他们保留或释放一个饲养员的可能性有多大,以及他们对捕鱼的兴趣会在多大程度上取决于他们以前的捕鱼成功。这也可以通过监测渔民并根据收集到的数据推断他们的行为来实现(Friedlander et al. 2014)。在其他地区,渔民离开渔场的意愿存在高度异质性,许多人即使失败也会继续捕鱼(Cinner et al. 2009, Muallil et al. 2011)。技能水平也会影响渔民的满意度。虽然数据的限制不允许我们包括渔民的技能水平属性,但未来的研究可以通过收集每个渔民钓鱼的年数和他们的捕鱼成功的数据来参数化这个变量。在其他地方,技术较差的渔民似乎更倾向于离开渔业,因为储量下降(van Poorten等人,2016)。虽然之前的成功以及技能水平可能是渔民捕鱼决策的重要组成部分,但该模型也可以受益于其他因素,如渔民的经济地位、天气和燃料价格(Cinner et al. 2009, Cooper and Jarre 2017, Daw et al. 2017)。尽管纳拉甘西特湾拥有大量的长期环境数据集,但它显然缺乏描述渔业属性的社会数据,而这些属性需要为渔民的决策提供细微差别。 Understanding the behavior of recreational fishers is essential to promote the sustainability and resilience of recreational fisheries (Arlinghaus and Cooke 2009, Arlinghaus et al. 2013)
管理的影响
我们的耦合模型的结果表明,包括人的维度,更具体地说,社会系统是如何表现的,即如何定义满意度损失等参数,可以改变模型的结果。这些发现为越来越多的文献提供了佐证,这些文献明确指出,需要将人类行为纳入环境,特别是渔业,以及使用基于生态系统的方法进行管理(即,Fulton等人2011年,Asah等人2014年,Hornborg等人2019年)。采取措施将渔民行为纳入管理的论文相对较少,它们往往侧重于经济指标(Hornborg等,2019年)。虽然经济刺激推动了商业渔业,但休闲渔业可能并非如此。例如,对渔人行为的研究表明,除了经济利益,符合社会规范的愿望、维护身份和经验尊重等因素也是重要的驱动因素(Hall-Arber et al. 2009)。事实上,渔民愿意为钓鱼的机会付费是很常见的(Cantrell et al. 2004, Johnston et al. 2006)。对于许多渔民来说,尽管经济损失仍继续捕鱼,捕鱼可以是一种娱乐方式,也是一种保护自我形象的方式(Wijermans等人,2020)。我们选择满意度作为我们的模型变量,因为它足够灵活,可以捕获这些非经济行为动机。在这项研究中,我们已经表明,结合休闲钓鱼动态可以导致不同的预测鱼类生物量和鱼的捕获数量,从而影响食物网。我们所演示的模型耦合框架对于管理战略评估特别有用,因为需要评估和平衡不同利益相关者冲突的目标(Smith et al. 1999)。 Going forward, it is imperative for scientists and managers to work with stakeholders to define the required metrics for assessing performance prior to conducting this research.
由于纳拉甘西特湾缺乏鱼类和渔民分布数据,我们没有将空间动态纳入食物网或渔民行为模型。饲料鱼的空间分布可能尤其重要,因为它们的分布比它们的数量更有可能影响它们的鱼食性捕食者(Hilborn et al. 2017)。如果这些数据是可用的,添加空间成分将增加现实主义和探索额外的社会生态问题,如在整个海湾不同地点的捕鱼成功率如何随着时间的推移而变化。空间分布还允许加入额外的行为因素,如渔民之间的竞争和合作,这在小规模渔业中尤其重要(Pollnac和Poggie 1991年,Basurto等人2016年)。我们模型中使用的种群年龄结构是由条纹鲈鱼数据估计的。如果有类似的蓝鲽鱼和夏季比目鱼数据,将有助于解决种群年龄结构对模型输出的影响。鱼类种群年龄结构是一个可以使用EwE模型的多节工具进一步探索的研究领域,该模型将功能群分成大小类别。此外,我们认为商业捕鱼压力是静态的,但在历史数据范围内。为了建立预测生态系统变化的模型,应将渔人行为模型扩展到包括商业船队动态及其对实时渔业盈利能力的响应。考虑到这些局限性,从我们的渔民行为模型中得出关于纳拉甘西特湾休闲渔业将如何随时间变化的具体结论不应该从这项工作中进行推断。 We aimed to create a reproducible yet flexible methodology for incorporating human behavior in a coupled social-ecological model built using established modeling platforms. We have also demonstrated that including these dynamics yields much different projections of important metrics such as fish biomass, catch, and fisher satisfaction. This model focuses on realism and generality as a goal, and accuracy can be further improved by filling the data gaps mentioned previously (Levins 1966).
考虑整个生态系统已成为渔业管理的一个高度优先事项,因为环境和人类的迅速变化已成为新常态;以生态系统为基础的渔业管理一直是全球渔业管理机构的最终目标,但迄今为止实施成功有限。可能是迄今为止最重要的应用之一,大西洋国家海洋渔业委员会采用了生态参考点来管理美国大西洋沿岸的大西洋鲱鱼,摆脱了单一物种的做法,现在通过在鲱鱼本身和捕食者(即条纹鲈鱼)的背景下管理鲱鱼的捕捞,制定了配额规范(SEDAR 2020)。然而,为了实现完全基于生态系统的渔业管理,这些生态系统方法必须通过模拟模型正式包括人类维度。在询问渔业权衡问题时,包括渔民行为尤其重要,例如与鲱鱼生态参考点相关的权衡(Chagaris等,2020年)。休闲渔民表示担心,鲱鱼捕捞正在耗尽他们喜欢的捕食者的猎物来源(Chase 2019)。在这样一个系统中,渔民行为的变化可能会对生态系统以及其他渔业部门产生深远的影响。我们已经证明,将渔民行为纳入模型可以改变鱼类生物量预测,因此应该是未来基于生态系统的渔业管理工作的优先事项。此外,我们还提供了创建这种耦合模型的方法。在这一点上,也许成功地将人类行为纳入模型的主要障碍是缺乏这些人类维度的数据。 In order to understand how ecosystems will change over time, researchers need to collect and incorporate data on the behavior of resource users. Although the absence of data on spatial distribution of fish and fishers, as well as region-specific fisher behavior, limits the predictive use of our social-ecological model, it is a useful tool for three main reasons. First, it has allowed us to explore general patterns of the linkage between the commercial and recreational fisheries of Narragansett Bay. Knowing the extent to which these two fishing sectors are linked is important when deciding if and how they should be considered in the management of one another. Second, our social-ecological model provides a base model to which new data can be added to expand the accuracy of the model’s representation of the Narragansett Bay recreational fishery. Finally, there are no examples of coupled EwE and agent-based models that we are aware of, making this method novel and providing a framework of how to incorporate human behavior for other fisheries to follow.
致谢
这项工作得到了罗德岛国家科学基金会建立的刺激竞争性研究资助项目#OIA-165522的支持。我们感谢Kelvin gorope、Tracey Dalton、Emi Uchida和Todd Guilfoos在模型的概念化和构建过程中提供的信息。我们还感谢国家社会环境综合中心(SESYNC)主办了一个基于代理的建模研讨会,以及两位匿名的审阅者提供了有用的反馈。这项研究是罗德岛海洋渔业研究所的一项贡献。本文所表达的观点仅代表作者本人,并不一定反映其所属机构的观点。
数据可用性
本研究中使用的模型可在EcoBase (http://ecobase.ecopath.org/php/protect/base_model.php?lang=&provenance=web&ident=&pass=&action=base&model=753)和CoMSES/OpenABM(基于代理的模型)。
文献引用
Arlinghaus, r . 2006。在休闲钓鱼的动机和满意度之间明显的脱节:德国垂钓者的渔获取向的例子。中国水产学报26(3):592-605。https://doi.org/10.1577/M04-220.1
Arlinghaus, R.和S. J. Cooke, 2009。休闲渔业:社会经济重要性、保护问题和管理挑战。B.迪克森,J.赫顿和W. M.亚当斯编辑的39-58页。休闲狩猎、保护和农村生计:科学与实践。布莱克威尔出版社,奇切斯特,英国https://doi.org/10.1002/9781444303179.ch3
Arlinghaus, R., S. J. Cooke和W. Potts, 2013。通过增进对鱼类和渔民行为的了解,在全球范围内发展有复原力的休闲渔业。渔业管理与生态20(2-3):91-98。https://doi.org/10.1111/fme.12027
Asah, s.t., a.d. Guerry, D. J. Blahna和J. J. Lawler. 2014。生态系统服务的感知、获取和使用:人类行为、生态系统管理和政策影响。生态系统服务10:180 - 186。https://doi.org/10.1016/j.ecoser.2014.08.003
大西洋国家海洋渔业委员会,2015。2015年大西洋条纹鲈鱼存量评估更新。大西洋州海洋渔业委员会,美国弗吉尼亚州阿灵顿。
Bannerot, S. P.和C. B. Austin, 1983。利用单位努力渔获量的频率分布来衡量鱼类种群丰度。美国渔业学会学报112(5):608-617。https://doi.org/10.1577/1548 - 8659 (1983) 112 < 608: UFDOCP > 2.0.CO; 2
Basurto, X., E. Blanco, M. Nenadovic和B. Vollan。将同时的亲社会和反社会行为纳入集体行动理论。科学进步2 (3):e1501220。https://doi.org/10.1126/sciadv.1501220
Boyd, J.和S. Banzhaf. 2007。什么是生态系统服务?需要标准化的环境核算单位。生态经济学63(2 - 3):616 - 626。https://doi.org/10.1016/j.ecolecon.2007.01.002
布赫斯特,A., T. J. Miller和E. D. Houde。评估大西洋鲱鱼的生态系统参考点。海洋和沿海渔业9(1):457-478。https://doi.org/10.1080/19425120.2017.1360420
伯吉斯、M. G、E. Carrella、M. Drexler、R. L. Axtell、R. M. Bailey、J. R. Watson、R. B. Cabral、M. Clemence、C. Costello、C. Dorsett、S. D. Gaines、E. S. Klein、P. Koralus、G. Leonard、S. A. Levin、L. R. Little、J. Lynham、J. K. Madsen、A. Merkl、B. Owashi、S. E. Saul、I. E. van Putten和S. Wilcox。在渔业的人类方面建立基于主体的模型的机会。渔业21(3):570-587。https://doi.org/10.1111/faf.12447
坎贝尔,2004年。利用一般线性模型建立空间变化渔业中CPUE的标准化和种群丰度指数。渔业研究70(2 - 3):209 - 227。https://doi.org/10.1016/j.fishres.2004.08.026
Cantrell, R. N., M. Garcia, P. Leung, D. Ziemann. 2004。康乐垂钓者愿意为增加的太平洋线鳍渔获率付费(Polydactylus sexfilis在夏威夷)。渔业研究68(1 - 3):149 - 158。https://doi.org/10.1016/j.fishres.2004.01.003
M. Cenek和M. Franklin, 2017年。在SES框架内指导多物种太平洋鲑鱼渔业管理的适应性基于代理的模型。生态造型360:132 - 149。https://doi.org/10.1016/j.ecolmodel.2017.06.024
Chagaris, D., K. Drew, A. Schueller, M. Cieri, J. Brito和A. Buchheister, 2020。利用中等复杂性生态系统模型建立大西洋鲱鱼的生态参考点。海洋科学前沿7:606417。https://doi.org/10.3389/fmars.2020.606417
Chagaris, D. D., B. Mahmoudi, C. J. Walters, M. S. Allen. 2015。利用Ecopath with Ecosim模拟渔业政策选择对西佛罗里达大陆架的营养影响。海洋与沿海渔业7(1):44-58。https://doi.org/10.1080/19425120.2014.966216
追逐,c . 2019。休闲渔业组织正式反对menhaden对地中海安全委员会的认可。海鲜资源新闻,3月29日。(在线)网址:https://www.seafoodsource.com/news/environment-sustainability/recreational-fishing-groups-formally-object-to-menhaden-s-msc-nod
克里斯滕森,V.和D.保利,1992。一个平衡稳态生态系统模型和计算网络特征的软件。生态模型61(3 - 4):169 - 185。https://doi.org/10.1016/0304 - 3800 (92) 90016 - 8
克里斯滕森,V., J. Steenbeek, P. Failler, 2011。综合生态系统和价值链建模方法,以评估捕捞的社会成本和效益。生态模型222(3):857 - 864。https://doi.org/10.1016/j.ecolmodel.2010.09.030
克里斯滕森,V., C.沃尔特斯和D.保利,2005。Ecopath with Ecosim:用户指南。加拿大温哥华英属哥伦比亚大学渔业中心和马来西亚槟城ICLARM。
辛纳,j。E。t。道和t。r。麦克拉纳汉。2009.影响手工渔民退出衰退渔业的社会经济因素。保护生物学23(1):124 - 130。https://doi.org/10.1111/j.1523-1739.2008.01041.x
Coll, M., A. Bundy, L. J. Shannon, 2009。生态系统建模使用Ecopath与Ecosim方法。b.a. Megrey和E. Moksness的225-291页,编辑。计算机用于渔业研究。荷兰,多德雷赫特,施普林格。https://doi.org/10.1007/978-1-4020-8636-6_8
柯利,J. S., A. D.伍德和H. P.杰弗里斯,2008。沿海鱼类群落物种组成的长期变化。加拿大渔业和水生科学杂志65:1352-1365。https://doi.org/10.1139/F08-048
1984年,j.g.库克和j.r.贝丁顿。渔场中渔获率与丰度之间的关系。数学医学与生物学1(4):391-405。https://doi.org/10.1093/imammb/1.4.391
R. Cooper和A. Jarre, 2017年。南非近海鳕拖网行业基于agent的模型:第一部分模型描述与验证。生态经济学142:268 - 281。https://doi.org/10.1016/j.ecolecon.2017.06.026
Cury, P., A. Bakun, R. J. M. Crawford, A. Jarre, R. A. Quiñones, L. J. Shannon和H. M. Verheye 2000。上升流系统中的小型远洋鱼类:“黄蜂腰”生态系统的相互作用模式和结构变化。海洋科学学报,57(3):603-618。https://doi.org/10.1006/jmsc.2000.0712
道尔顿,T., R.汤普森和D.金,2010。海洋空间规划和管理中的人类维度映射:罗德岛纳拉甘塞特湾的例子。海洋政策34(2):309 - 319。https://doi.org/10.1016/j.marpol.2009.08.001
道,T. M., J. E. Cinner, T. R. McClanahan, K. Brown, S. M. Stead, N. A. J. Graham和J. Maina. 2017。捕鱼或不捕鱼:影响手工渔民准备退出衰退渔业的多种尺度因素。12 (2): PLOS ONE e0172075。https://doi.org/10.1371/journal.pone.0172075
迪奥达蒂,P. J.和R. A.理查兹,1996。将条纹鲈鱼钩入海水放生后的死亡率。美国渔业学会学报125(2):300-307。https://doi.org/10.1577/1548 - 8659 (1996) 125 < 0300: MOSBHA > 2.3.CO; 2
恩格尔哈德、g.h.、m.a.派克、A.林多夫、S. C.斯莫特、M. van Deurs、K. Raab、K. H. Andersen、S. Garthe、R. A. M. Lauerburg、F. Scott、T. Brunel、G. Aarts、T. van Kooten和M. Dickey-Collas。2014.饲料鱼,它们的渔场,它们的掠食者:谁赶谁?海洋学报71(1):90- 99。https://doi.org/10.1093/icesjms/fst087
埃辛顿,T. E., P. E.莫里亚蒂,H. E. Froehlich, E. E. Hodgson, L. E. Koehn, K. L. Oken, M. C. simple和C. C. Stawitz. 2015。捕鱼加剧了饲料鱼种群的减少。美国国家科学院学报112(21):6648-6652。https://doi.org/10.1073/pnas.1422020112
Friedlander, A., P. Curry, M. Donovan, A. Filous, J. Giddens, W. Goodell, K. Kamikawa, H. Koike, E. Schemmel, K. Stamoulis和P. Usseglio. 2014。评估瓦胡岛迎风方向受保护和未受保护区域的布施刺网法规。渔业生态研究实验室,夏威夷大学,檀香山,夏威夷,美国。
弗罗斯,R.和D.保利,2019年。FishBase。https://www.fishbase.org
富尔顿,e.a., a.d.m.史密斯,d.c.史密斯和i.e.m van Putten, 2011。人类行为:渔业管理不确定性的主要来源。鱼类及渔业12(1):2-17。https://doi.org/10.1111/j.1467-2979.2010.00371.x
和M. Dreyfus-Leon。2004.用人工神经网络模拟金枪鱼围网渔业单位努力渔获量与丰度之间的非线性关系。海洋学报61(5):812-820。https://doi.org/10.1016/j.icesjms.2004.05.002
格林、V. U.伯杰、F.巴斯蒂安森、S.埃利亚森、V.吉诺、J.吉斯克、J.戈斯-卡斯塔德、T.格兰德、S. K.海因茨、G.豪斯、A.胡特、J. U.杰普森、C. J. Jørgensen、W. M. Mooij、B. Müller、G.佩尔、C. Piou、S. F. Railsback、A. M. Robbins、M. M. Robbins、E. rosmanith、N. Rüger、E. Strand、S. Souissi、R. A. Stillman、R. Vabø、U. Visser、D. L. DeAngelis。2006.描述基于个体和基于代理模型的标准协议。生态模型198(1):115 - 126。https://doi.org/10.1016/j.ecolmodel.2006.04.023
格林,V., U.伯杰,D. L. DeAngelis, J. G. Polhill, J. Giske, S. F. Railsback, 2010。ODD协议:回顾和首次更新。生态模型221(23):2760 - 2768。https://doi.org/10.1016/j.ecolmodel.2010.08.019
格林,V. S. F. Railsback, C. E. Vincenot, U. Berger, C. Gallagher, D. L. DeAngelis, B. Edmonds, J. Ge, J. Giske, J. Groeneveld, A. S. A. Johnston, A. Milles, J. Nabe-Nielsen, J. G. Polhill, V. Radchuk, m.s。Rohwäder、r.a. Stillman、j.c. Thiele和D. Ayllón。2020.用于描述基于代理和其他模拟模型的ODD协议:第二次更新,以提高清晰度、复制性和结构逼真性。人工社会与社会模拟23(2):7。https://doi.org/10.18564/jasss.4259
格雷罗,A. M., N. J. Bennett, K. A. Wilson, N. Carter, D. Gill, M. Mills, C. D. Ives, M. J. Selinske, C. Larrosa, S. Bekessy, F. A. Januchowski-Hartley, H. Travers, C. A. Wyborn和A. Nuno。实现社会生态研究一体化的前景:综述与展望。生态与社会23(3):38。https://doi.org/10.5751/ES-10232-230338
1969年,j.a.古兰。第五节。死亡率。鱼类资源评估方法手册-第1部分。鱼人口分析。联合国粮食及农业组织,罗马,意大利。
哈格蒂和J. R.金,2006。ccpu作为近岸礁鱼相对丰度的指标。渔业研究81(1):89 - 93。https://doi.org/10.1016/j.fishres.2006.05.015
Hall-Arber, M., C. Pomeroy和F. Conway, 2009。计算渔业的人类维度:具有启发性的模型。海洋和沿海渔业1(1):300-314。https://doi.org/10.1577/C09-006.1
Hangsleben, m.a., m.s. Allen, D. Gwinn, 2013。衡量佛罗里达湖泊鱼类丰度的单位努力电捕渔获量评价。美国渔业学会学报142(1):247-256。https://doi.org/10.1080/00028487.2012.730106
汉森,a.g. 2018年。冷水湖泊和水库研究项目。年度报告,科罗拉多公园和野生动物,柯林斯堡,科罗拉多,美国。
哈利,S. J., R. A.迈尔斯和A.邓恩,2001年。单位努力捕获量与丰度成比例吗?加拿大水产科学杂志58(9):1760-1772。https://doi.org/10.1139/f01-112
海曼斯,J. J., M. Coll, J. S.林克,S. Mackinson, J. Steenbeek, C. Walters和V. Christensen. 2016。Ecosim食品网络模型的生态医疗的最佳实践基于生态系统的管理。生态造型331:173 - 184。https://doi.org/10.1016/j.ecolmodel.2015.12.007
Hilborn, r . 1985。船队动态和个体变化:为什么有些人比其他人捕获更多的鱼。加拿大水产科学杂志42:2-13。https://doi.org/10.1139/f85-001
Hilborn, R., R. O. Amoroso, E. Bogazzi, O. P. Jensen, A. M. Parma, C. Szuwalski, C. J. Walters. 2017。捕鱼的饲料品种什么时候影响它们的捕食者?渔业研究191:211 - 221。https://doi.org/10.1016/j.fishres.2017.01.008
Hornborg, S., I. van Putten, C. Novaglio, E. A. Fulton, J. L. Blanchard, É。Plagányi, C. Bulman和K. Sainsbury, 2019年。基于生态系统的渔业管理需要更广泛的人类维度绩效指标。海洋政策108:103639。https://doi.org/10.1016/j.marpol.2019.103639
Houle, J. E., K. H. Andersen, K. D. Farnsworth和D. G. Reid。2013。草料渔业和掠食渔业之间出现的不对称相互作用造成了管理上的权衡。水生生物学报83(4):890-904。https://doi.org/10.1111/jfb.12163
Innes-Gold, A., M. Heinichen, K. Gorospe, C. Truesdale, J. Collie和A. Humphries。模拟纳拉甘西特湾25年的食物网变化,作为基于生态系统的管理工具。海洋生态进展系列654:17-33。https://doi.org/10.3354/meps13505
约翰斯顿,R., M.兰德森,E.贝西丁和E.赫尔姆,2006。是什么决定了每条鱼的支付意愿?休闲钓鱼价值的元分析。海洋资源经济21(1):1-32。https://doi.org/10.1086/mre.21.1.42629492
Kaemingk, m.a., K. L. Hurley, C. J. Chizinski, K. L. Pope, 2020。休闲渔业的放收决策。中国水产科学学报,29(3):329 - 336。https://doi.org/10.1139/cjfas-2019-0119
基默勒,W., S. R. Avent, S. M. Bollens, F. Feyrer, L. F. Grimaldo, P. B. Moyle, M. Nobriga, T. Visintainer, 2005。根据调查数据估算河口鱼类生物量所用的长度-重量关系的变异性。美国渔业学会学报134(2):481-495。https://doi.org/10.1577/T04-042.1
克雷布斯,C. J. 2014。存活率的估计。生态方法论第655-701页。第三版,在准备。艾迪生韦斯利,波士顿,马萨诸塞州,美国。
Kuffner, a . 2017。呼吁采取新办法管理鲱鱼渔业。10月5日《普罗维登斯杂志》
大型,1992年。利用乘法模型估计商业CPUE数据的相对丰度。海洋学报49(3):253-262。https://doi.org/10.1093/icesjms/49.3.253
莱文,r . 1966。种群生物学中的模型建立策略。美国科学家54(4):421 - 431。
刘,J., T. Dietz, S. R. Carpenter, M. Alberti, C. Folke, E. Moran, A. N. Pell, P. Deadman, T. Kratz, J. Lubchenco, E. Ostrom, Z. Ouyang, W. Provencher, C. L. Redman, S. H. Schneider和W. W. Taylor。2007。人类和自然系统耦合的复杂性。科学》317(5844):1513 - 1516。https://doi.org/10.1126/science.1144004
露西,J.和T. D.霍尔顿,1998。在维吉尼亚休闲渔场释放夏季比目鱼牙鲆的死亡率。海洋资源报告第97-8号;vsg - 97 - 09年。威廉玛丽学院弗吉尼亚海洋科学研究所,格洛斯特点,美国弗吉尼亚州。https://doi.org/10.21220/m2-e77c-5m84
麦卡尔,C. M.和M. J.诺斯,2005。基于代理的建模和仿真教程。冬季模拟会议论文集。电气和电子工程师学会,皮斯卡塔韦,美国新泽西州。https://doi.org/10.1109/WSC.2005.1574234
麦凯,M.,山崎S., J. M. Lyle和E. M. Ogier, 2020。确定多方法消费休闲渔业的管理偏好。生态学报25(2):22。https://doi.org/10.5751/es-11602-250222
M. H.马尔科夫和J. A.露西1998。纽约和弗吉尼亚夏季比目鱼的短期钩死率。海洋资源报告第98-7号。威廉玛丽学院弗吉尼亚海洋科学研究所,格洛斯特点,美国弗吉尼亚州。https://doi.org/10.21220/m2-mw7g-bj76
S. J. D., A. I. Beattie, C. J. Walters, T. Nayar和R. Briese 2002。利用Ecopath和Ecosim软件模拟格鲁吉亚海峡生态系统渔业管理策略。渔业中心研究报告10(2):16-23。
Martino, E. J.和E. D. Houde, 2012。0岁条纹鲈鱼幼鱼年级强度的密度依赖性调节(马龙saxatilis).中国水产科学,29(3):429 - 436。https://doi.org/10.1139/f2011-149
2011年,马特奥,德宾,本特森和杜兰特。纳拉甘西特湾苗圃和罗德岛沿岸池塘中陶陶克的生长变化。海洋及沿岸渔业3(1):271-278。https://doi.org/10.1080/19425120.2011.601943
Matsumura, S., B. Beardmore, W. Haider, U. Dieckmann, R. Arlinghaus. 2019。生态、垂钓者和空间异质性驱动淡水休闲渔业综合景观模型的社会和生态结果。水产科学进展27(2):170-197。https://doi.org/10.1080/23308249.2018.1540549
孟,L., C. D. Orphanides和J. C. Powell 2002。使用鱼类指数来评估罗德岛纳拉甘西特湾的栖息地质量。美国渔业学会学报131(4):731-742。https://doi.org/10.1577/1548 - 8659 (2002) 131 < 0731: UOAFIT > 2.0.CO; 2
2018年,美瑟尔和斯普劳尔。罗得岛州渔业和海产品的经济影响。商业渔业研究基金会,金斯顿,罗德岛,美国。
莫尔斯,1981年。夏天比目鱼的繁殖,Paralichthys dentatus(l)。鱼类学报19(2):189-203。https://doi.org/10.1111/j.1095-8649.1981.tb05823.x
穆阿利尔,R. N., R. C.杰罗尼莫,D.克莱兰,R. B.卡布拉尔,M. V.博士,A.克鲁兹-特立尼达,P. M. Aliño。2011.愿意退出手工渔业,以应对捕鱼量下降或货币激励增加的情况。渔业研究111(1):74 - 81。https://doi.org/10.1016/j.fishres.2011.06.013
纳拉甘塞特湾入海口项目。纳拉甘西特湾州及其分水岭。技术报告。纳拉甘西特湾河口项目,普罗维登斯,罗德岛,美国。
国家海洋渔业局,2018。2016年美国渔业经济报告。美国国家海洋和大气管理局技术备忘录。NMFS-F / SPO-187:243。美国商务部,华盛顿特区,美国。
美国国家海洋和大气管理局(NOAA)渔业。夏天的比目鱼。NOAA渔业中心,华盛顿特区,美国。新泽西州鱼类和野生动物部,特伦顿,新泽西州,美国。(在线)网址:https://www.fisheries.noaa.gov/species/summer-flounder
美国国家海洋和大气管理局(NOAA)渔业。蓝。NOAA渔业中心,华盛顿特区,美国。https://www.fisheries.noaa.gov/species/bluefish
美国国家海洋和大气管理局(NOAA)渔业。大西洋条纹鲈鱼。NOAA渔业中心,华盛顿特区,美国。https://www.fisheries.noaa.gov/species/atlantic-striped-bass
纳尔逊,1998年。北卡罗来纳州罗阿诺克河条纹鲈鱼的捕获和释放死亡率。北美渔业管理杂志18(1):25-30。https://doi.org/10.1577/1548 - 8675(1998) 018 < 0025:•> 2.0.CO; 2
新泽西鱼类和野生动物部,2010。条纹低音长度/年龄频率数据。新泽西州鱼类和野生动物部,特伦顿,新泽西州,美国。(在线)网址:https://www.state.nj.us/dep/fgw/artstrpbass10_pbsample.htm
尼克松,S. W., B. A.巴克利,S. L.格兰杰,L. A.哈里斯,A. J. Oczkowski, R. W. Fulweiler和L. W. Cole。2008。纳拉甘西特湾氮、磷输入:过去、现在和未来。编辑:A. Desbonnet和B. A. Costa-Pierce,第101-175页。基于生态系统的管理科学:21世纪的纳拉甘西特湾。施普林格,纽约,美国纽约。https://doi.org/10.1007/978-0-387-35299-2_5
诺布丽加,M. L.和F.费勒,2008。旧金山河口条纹鲈鱼的饮食组成:营养适应性有其局限性吗?鱼类环境生物学83(4):495-503。https://doi.org/10.1007/s10641-008-9376-0
诺兰,J., D. Parker, G. C. van Kooten和T. Berger。2009。农业与资源经济学的计算模型综述。加拿大农业经济杂志/加拿大农业经济杂志57(4):417-429。https://doi.org/10.1111/j.1744-7976.2009.01163.x
Ono, K., A. Haynie, A. Hollowed, J. Ianelli, C. McGilliard, and A. Punt. 2017。多物种渔业的管理策略分析,包括管理决策和捕鱼建模中的技术交互和人类行为。中国水产科学,29(5):362 - 362。https://doi.org/10.1139/cjfas-2017-0135
Oviatt, C., S. Olsen, M. Andrews, J. Collie, T. Lynch, K. Raposa. 2003。纳拉甘西特湾一个世纪以来的渔业和鱼类波动。水产学报11(3):221-242。https://doi.org/10.1080/10641260390244413
Oviatt, C., L. Smith, J. Krumholz, C. Coupland, H. Stoffel, A. Keller, M. C. McManus, L. Reed. 2017。在北温带河口,有管理的营养物减少对营养物浓度、水透明度、初级生产量和缺氧的影响。海洋科学,1999:25- 25。https://doi.org/10.1016/j.ecss.2017.09.026
帕洛海莫,j.e.和l.m.迪基,1964年。富足和捕鱼的成功。联系Procès-verbaux des Réunions du Conseil International pour Exploration de la Mer 155:152-163。
Pascoe, S., T. Cannard, E. Jebreen, C. M. Dichmont和J. Schirmer. 2015。钓鱼的满足感和离开的欲望。中44(5):401 - 411。https://doi.org/10.1007/s13280-014-0579-7
保利,D., V.克里斯滕森和C.沃尔特斯,2000。Ecopath、Ecosim和Ecospace作为评估渔业对生态系统影响的工具。海洋科学学报,57(3):697- 697。https://doi.org/10.1006/jmsc.2000.0726
彼得曼,R. M.和G. J.斯蒂尔,1981。运动捕鱼捕集系数与鲑鱼丰度的关系。美国渔业学会学报110(5):585-593。https://doi.org/10.1577/1548 - 8659 (1981) 110 < 585: RBSCCA > 2.0.CO; 2
皮尔,L., S. P.巴登,R. J.迪亚兹和L. C.沙夫纳,1992。低氧引起底栖鱼类和甲壳类食物结构的变化。海洋生物112(3):349 - 361。https://doi.org/10.1007/BF00356279
Pitman、K. J.、S. M. Wilson、E. Sweeney-Bergen、P. Hirshfield、M. C. Beere和J. W. Moore。将垂钓者,鱼和管理联系起来,在一个捕获和释放虹鳟渔业。加拿大渔业和水生科学杂志=加拿大海洋科学杂志76(7):1060-1072。https://doi.org/10.1139/cjfas-2018-0080
波纳克,r.b.和J. J.波吉1991。厄瓜多尔小型渔民合作社的心理文化适应和发展政策。人类组织50(1):43-49。https://doi.org/10.17730/humo.50.1.97tm3638277868n7
1984年,j.j.波洛维娜。珊瑚礁生态系统模型。珊瑚礁3(1):1 - 11。https://doi.org/10.1007/BF00306135
罗德岛州的民主党。2020.海洋渔业的最小规模和拥有限制。罗德岛DEM,普罗维登斯,罗德岛,美国。(在线)网址:http://www.dem.ri.gov/programs/marine-fisheries/mfsizes.php
理查兹,L. J.和J. T.施纽特,1986。对这个问题的实验和统计方法:CPUE是丰度指数吗?中国水产科学,29(6):1214-1227。https://doi.org/10.1139/f86-151
罗毕拉德,E., C. S. Reiss和C. M. Jones, 2008。蓝鱼的繁殖生物学(Pomatomus saltatrix)沿美国东海岸。渔业研究90(1 - 3):198 - 208。https://doi.org/10.1016/j.fishres.2007.10.020
Schlüter, M., R. R. J. Mcallister, R. Arlinghaus, N. Bunnefeld, K. Eisenack, F. Hölker, E. J. Milner‐Gulland, B. Müller, E. Nicholson, M. Quaas, M. Stöven。2012.环境管理的新视野:耦合社会-生态系统模型综述。自然资源学报25(1):219-272。https://doi.org/10.1111/j.1939-7445.2011.00108.x
谢弗,j.l., 2007。基于智能体的休闲珊瑚礁渔业模拟:连接社会和生态动态。博士论文,夏威夷大学马诺阿,夏威夷,美国。
Sharov, A., G. Nelson, H. Corbett, G. Wippelhauser, K. Sullivan, G. Shepherd, N. Lengyel, K. Gottschall, A. Kahnle, K. Hattala, K. McShane和C. Hoffman。2013。2013年条纹鲈鱼股票评估。页492 - 967。第57届东北区域储量评估研讨会(第57届SAW)评估报告。东北渔业科学中心,伍兹霍尔,马萨诸塞州,美国。
A. D. M. Smith, K. J. Sainsbury和R. A. Stevens. 1999。实施有效的渔业管理系统-管理战略评估和澳大利亚伙伴关系办法。海洋学报56(6):967-979。https://doi.org/10.1006/jmsc.1999.0540
东南数据,评估和审查(SEDAR)。2015.SEDAR 40 -大西洋鲱鱼种群评估报告。西达,北查尔斯顿,南卡罗莱纳州,美国。
东南数据,评估和审查(SEDAR)。2020.生态参考点种群评估报告:大西洋鲱鱼。西达,北查尔斯顿,南卡罗莱纳州,美国。
Szczepanski, j . 2013。德拉瓦湾和纳拉甘西特湾鳐鱼的摄食生态。论文。罗德岛大学,金斯顿,罗德岛,美国。https://doi.org/10.23860/diss-szczepanski-john-2013
Terceiro m . 2010。2010年夏季鲽鱼种群评估。美国国家海洋和大气管理局渔业中心,东北渔业科学中心,伍兹霍尔,马萨诸塞州,美国。
Tsuboi, J.和S. Endou. 2008。根据山溪中鲑鱼的实际测量,单位努力渔获量、可捕性和丰度之间的关系。美国渔业学会学报137(2):496-502。https://doi.org/10.1577/T06-259.1
van Poorten, B. T., C. J. Walters和H. G. M. Ward, 2016。通过努力分类预测渔获力系数的变化,因为技术较差的渔民在渔获量下降时退出渔业。渔业研究183:379 - 384。https://doi.org/10.1016/j.fishres.2016.06.023
怀特,j.w., A.拉斯韦勒,J. F. Samhouri, A. C.斯蒂尔和C.怀特。2014。生态学家不应该使用统计显著性检验来解释模拟模型的结果。Oikos 123(4): 385 - 388。https://doi.org/10.1111/j.1600-0706.2013.01073.x
Wijermans, N., W. J. Boonstra, K. Orach, J. Hentati‐Sundberg和M. Schlüter。2020.捕鱼行为多样性-迈向下一代渔业模式。渔业科学,21(5):872-890。https://doi.org/10.1111/faf.12466
Wildenhain, e . 2016。大口黑鲈的捕鱼能力(Micropterus salmoides)生长在美国北部一个温带湖泊的粗木栖地。论文。诺特丹大学环境研究中心,Land O 'Lakes,威斯康星州,美国。(在线)网址:https://underc.nd.edu/assets/215459/fullsize/wildenhain2016.pdf
奥巴马,美国1999年。NetLogo。连接学习和计算机建模中心,西北大学,伊文森,伊利诺斯州,美国。
