






以下是引用本文的既定格式:
斯摩霍恩-韦斯特,P. F., P. J. Cohen, R. A. Morais, F. A. Januchowski-Hartley, D. Ceccarelli, S. Malimali, K. Stone, R. Warren和J. E. Cinner。部分保护珊瑚礁渔业的潜在利益和风险。生态与社会27(1):26。摘要
部分保护区现在是全球空间管理的主要形式,旨在保护生态系统的完整性和管理人类使用。然而,大多数对其有效性的评估只使用了一套狭窄的保护指标,这些指标反映了保护成功或失败的一小部分方式。在本文中,我们研究了三个部分保护的珊瑚礁渔业系统的案例,以评估其作为管理工具的效益和风险。我们利用来自三个太平洋岛屿国家的社区管理安排的数据,展示了三个小故事,说明部分保护如何能够促进渔业生产,提高渔民捕获猎物的难度,并改变渔业产量的构成。渔业生产力、捕获量和部分保护下的脆弱性的这些变化为渔民带来了巨大的利益。然而,它们也对生态系统和渔业生计构成重大风险,除非进行适应性管理,以便在不危及长期可持续性的情况下在资源表现方面带来短期和中期效益。介绍
21世纪的一个关键需求是平衡自然保护和可持续利用(Halpern et al. 2013)。保护区限制资源开采已经成为陆地、海洋和内陆水生系统管理的流行工具(Joppa和Pfaff 2011, Edgar等人2014,Visconti等人2015)。保护区的使用继续扩大,建议的目标是到2030年将地球上30%的区域置于某种形式的空间管理之下(Waldron et al. 2020)。然而,在保护区内的控制和限制在资源的使用受到限制的程度和对象上可能有所不同。尽管随着时间的推移,完全关闭(禁止开采活动,甚至禁止进入)可以为生物多样性保护带来实质性的好处(Joppa和Pfaff 2011, Edgar et al. 2014),但它们也可能带来巨大的成本。短期内,在人们的生计和粮食安全依赖于边界内资源的地区,这些成本可能特别高(Ferraro和Hanauer 2011年,Brockington和Schmidt-Soltau 2017年)。因此,在许多情况下,完全将某一地区的人排除在外并完全禁止使用他们所依赖的资源在政治上和经济上都不可行,并可能导致巨大的社会经济成本(Halpern et al. 2013)。
尽管考虑到一整套可用的管理工具是可取的,但在许多情况下,利益相关者的目标和价值将把某些工具排除在解决方案空间之外(McClanahan等人2012,Haasnoot等人2020)。部分保护区(PPAs)允许在边界内进行有限的资源开采,是一种广泛应用的措施,它规定了何时、如何以及由谁进行采收(Jupiter等人2014年,Zupan等人2018年,Turnbull等人2021年)。部分保护区现在构成了全球海洋空间管理的主要形式(占所有海洋管理区的69%)(Turnbull等人,2021年)。然而,作为一种管理工具,PPAs的价值取决于期望的结果是源自保护还是粮食安全和生计,而且PPAs带来的生态效益有多种证据(Lester和Halpern 2008, Giakoumi等人2017,Sala和Giakoumi 2018, Zupan等人2018,Turnbull等人2021)。例如,在渔业领域,部分保护已被证明可以同时改善鱼类生物量、鹦嘴鱼放牧和性状多样性,但仅在特定的社会经济背景下,其程度远低于全面保护(Cinner等,2020年)。部分保护可以带来重大的生计效益,例如,如果定期打开封地进行收割,可提高封地内的收割效率(Cohen和Alexander 2013年,Goetze等人2017年,Carvalho等人2019年,Cinner等人2019年)。然而,如果管理不当,收获效率的提高也会导致过度收获(Robinson et al. 2015, Goetze et al. 2017)。
实施ppa的治理结构的差异也可能导致不同的结果。例如,与自上而下的系统制定相同的规则相比,在自下而上或协作管理(通常称为共同管理)系统中集体制定部分保护规则更有可能获得好处,这些系统往往获得较高的合法性和合规性(Ban et al. 2019)。
在这里,我们用共同管理的珊瑚礁系统的镜头来说明在这些制度下部分保护渔业的好处和风险。通过对太平洋岛屿国家的三个案例研究,我们评估了PPAs的三个渔业指标——潜在渔业生产力、个体鱼类捕获量和内在脆弱性——与公开捕鱼区和/或充分保护的区域的比较。我们研究:(1)汤加的渔业生产力潜力;(2)瓦努阿图的个体捕鱼量;(3)所罗门群岛的内在渔获脆弱性(图1;表1).我们注意到检查这三个案例并没有反映一个完全交叉的样本设计;每一情况的数据只适合评估现有的一项指标。然而,每个案例的研究结果都代表了PPAs如何影响渔业效益,因此,尽管在跨其他系统推断这些研究结果方面存在局限性(对个别案例的大多数研究也出现这种情况),但结合起来研究是有用的。我们还注意到,许多海洋管理区明确以保护成果和维持生态功能为目标,而我们的综合研究则侧重于渔业成果。然而,这些案例研究——其中两个讨论了整个管理领域网络的发现(汤加和瓦努阿图分别是生产力和可获取性),一个讨论了十年监测的结果(所罗门群岛的脆弱性)——提供了一系列小故事,说明了部分保护对可持续资源管理可能带来的关键好处和风险。
合成
生产力
潜在渔业生产力(以下简称”生产力”(kg ha(¹day)¹)表示系统中目标鱼的生物量累积率,根据观察到的个体的种类和大小,根据其预期的生长和存活情况(Morais et al. 2020)。这与渔业产量和生物量积累(MacNeil et al. 2015)相比,提供了更直接的联系,因为生物量是一个静态的衡量指标,由产量在一个未知的时间框架内累积产生。因此,生产力是高度多样性热带生态系统中目标鱼类组合的潜在渔业产量(即可以收获什么)的独立指标。生产力被计算为两个相反的动态平衡作用于单个鱼:体细胞生长和死亡。第一个组成部分,日生长量,由von Bertalanffy生长模型从形态和行为特征(最大物种体型、饮食、相对于礁石的位置和海洋表面温度)预测的轨迹量化。然后,通过概率程序纳入第二个组成部分,即死亡率,根据根据物种、个体体型和海洋表面温度估算的预期瞬时死亡率,将命运(存活率vs.死亡率)分配给单个鱼类(Morais等,2019年)。关于计算渔业潜在生产力的详细方法,请参阅Morais和Bellwood(2020年)。
虽然计算渔业产量并不是一个新概念(例如,Brock 1985, Welcomme and Bartley 1998, Finney et al. 2002),但一个容易量化的多物种珊瑚礁系统生产力指标直到最近才出现。Morais和Bellwood(2020)提供了详细的框架和R包(rfishprod),以量化多样性高的珊瑚礁的潜在渔业生产力,所用的数据与常用来量化直立鱼类生物量的水下目视调查数据相同。到目前为止,研究已经揭示了常存鱼类生物量和潜在渔业生产力之间的关系并不是紧密耦合的,因此,生物量准确代表潜在渔业产量的假设可能并不成立(Morais等人,2020年)。常设鱼类生物量和潜在渔业生产力对开发率的反应不同:当尺寸选择性开发耗尽鱼类生物量时,它会触发单位生物量产量的增加,这在模型和经验系统中都避免了立即的生产力崩溃。这一过程,称为”缓冲效率,”证明了使用生物量估算作为渔业潜力的替代的危险,也可能有助于解释为什么一些耗尽了生物量的珊瑚礁仍然可以支持活跃的渔业(Jennings和polun1996年,Morais和Bellwood 2020年)。在全球范围内,礁鱼功能类群之间的生产力估计也存在显著差异,礁鱼组合的大部分生产力来自于浮游食性物种和水柱光合作用,而不是封闭的礁鱼能量通道(Morais等人2019,2021)。重要的是,这些高比例的浮游食性生产力也可以在受影响的低珊瑚覆盖的珊瑚礁上维持,这为遭受珊瑚损失的珊瑚礁系统提供了希望,即如果管理得当,仍然可以维持较高的鱼类生产力,从而维持活跃的渔业。
Catchability
实施周期性捕获PPAs的一个关键原理是改变鱼类的行为,使它们更容易接近,从而更容易捕获(Foale et al. 2011)。个体渔获力(下同”catchability”)的定义是指渔民在不受鱼群密度影响的情况下捕获鱼类的难易程度,并根据渔民在鱼叉距离内接近目标礁鱼的概率来计算。通过对渔民的采访,可以确定鱼叉的距离。在采访中,渔民被要求估计鱼叉枪的最大有效射程,或者他们需要离鱼多近才能成功地鱼叉(Januchowski-Hartley et al. 2014)。然后,利用飞行起始距离(FID),根据鱼警惕性的行为调查(手拿长矛在水下接近鱼)来计算Fisher接近距离。鱼叉距离可以根据捕鱼者的经验和装备质量,以及特定环境的因素,如海洋条件而变化。高(或长)FID加上短的叉距表明鱼的捕获能力较低,而低(或短)FID加上高的叉距则表明鱼的捕获能力较高。
尽管管理和鱼类警惕之间的关系已经得到了充分的记录(例如Januchowski-Hartley等人,2012,2013,2014,2015,Goetze等人,2017),这在多大程度上影响了渔民的实际捕获概率,也没有量化这是如何受大小选择性的影响。鱼类警觉度的变化受到海洋管理的强烈影响,FID通常随着管理的实施而迅速下降,比生物量积累等其他影响更快,然后在重大收获事件后再次迅速上升(Januchowski-Hartley等,2014)。这种动态反应可能是土著人民和当地社区几个世纪以来一直使用周期性收获的PPAs来管理其水资源的一个关键原因(Cohen和Steenbergen, 2015)。这些变化也发生在广阔的空间尺度上,并沿着开发梯度发生,随着渔网捕捞压力水平的增加,鱼类的警觉性不仅在禁捕保护区之外,而且在禁捕保护区之内(Januchowski-Hartley et al. 2015)。鱼类naïveté也可以从管理区域溢出到周围的渔区,并比其他变量(如站立生物量)的变化保持更大的距离(Januchowski-Hartley et al. 2013)。此外,目标鱼种(Sbragaglia et al. 2018)和非目标鱼种(Tran et al. 2016)也可以很容易地区分鱼叉,这表明鱼类可以形成良好的反捕食者反应,并识别鱼叉捕鱼者作为捕食者带来的风险。
脆弱性
对捕捞的内在脆弱性被定义为一个物种承受剥削所造成的额外死亡率的内在能力,主要由种群的内在增长率(r) (Abesamis et al. 2014)。重要的是,内在脆弱性不同于外在脆弱性,外在脆弱性是基于暴露于风险的模式。例如,在本文中,PPAs的使用、禁捕封闭和公开捕鱼区域代表了外在脆弱性水平的差异,而物种关键生活史特征的差异则代表了它们内在的脆弱性(以下简称脆弱性)”脆弱性”)到过度捕捞。张等人(2007)利用模糊逻辑专家系统,根据8个生命史和生态特征(最长体长、首次成熟年龄、寿命、生长率、自然死亡率、繁殖力、空间行为和地理范围),对1353种鱼类和87个科(等级为1-100,其中100最容易被过度开发)制定了脆弱性指数。Abesamis等人(2014)用同样的方法又增加了145种珊瑚礁鱼类,它们属于10个科。输入变量由被认为与物种内在脆弱性相关的性状组成,这些性状来自一篇文献综述(张et al. 2005)。简单地说,专家系统以不同的关联程度将鱼类分类为不同的生活史类别,范围从0(无关联)到1(完全关联),并由预先定义的模糊逻辑隶属函数确定(张et al. 2005)。采用IF(谓词)- THEN(结论)子句表达的语言规则来推断物种的脆弱性水平(如IF最大长度大,则内在脆弱性高)。这一复合指标提供了一种简单的方法来描述多物种渔业开发的总体风险,平均值的下降表明渔获物中脆弱物种(例如生长缓慢和繁殖力低的物种)减少。
自1950年以来,全球捕鱼量越来越多地由内在脆弱性低的物种主导(张et al. 2007)。这种模式在珊瑚礁上表现得最为明显,2005年,珊瑚礁中脆弱的物种数量比1950年减少了25%。脆弱性也与广泛的数据集和生态系统中观察到的种群下降率显著相关,过度捕捞持续导致整体脆弱性得分下降,因为更脆弱的物种(即得分较高的物种)被过度捕捞,不再存在于系统中(张et al. 2005)。这些趋势也被用于预测物种恢复的模式,高度脆弱的物种比不那么脆弱的物种需要更长的时间(即几十年)来达到当地的承载能力(Abesamis等人,2014)。
材料和方法
案例研究1:汤加的生产力
2002年,汤加建立了特别管理区(SMA)计划,这是汤加社区和渔业部之间的一种协作渔业管理方法(Smallhorn-West等,2020年)一个).它代表了一种由两部分组成的海洋管理方法,即社区被授予对其社区(即PPA)周围珊瑚礁区域的独家使用权,作为交换,其中的一部分将成为禁止捕捞的保护区。第一个特别管理区于2006年实施,该计划目前包括>50社区的100个禁止进入或限制进入的PPAs。我们的研究考察了SMA计划对礁鱼生产力和生物量模式的影响(图1)。具体地说,我们是建立在之前的研究的基础上的,这些研究已经探索了常设生物量、密度、大小以及整体物种多样性(Smallhorn-West等,2020年)b),以计算目标鱼类在公开捕捞、限制进入的ppa区和完全封闭的地区的生物量(即潜在渔业生产力)的产生率。我们的影响评估只涵盖2014年之前建立的管理区域,以及生态调查时至少3年的管理区域。因此,在这项研究中,我们纳入了七个社区,每个社区都实施了一个禁止捕捞保护区和一个限制准入PPA,只有该社区的成员可以在那里捕鱼。禁采储量为0.5 ~ 2 km²,禁采储量为5 ~ 40 km²。
2016年至2018年,在汤加的375个地点进行了生态调查,包括管理区内(7个禁止进入和7个限制进入)和潜在的控制地点。在每个地点,在3-12米(捕鱼最常见的深度)的深度平行于珊瑚礁等高线布设了4至6条30米的样带(共1635条),在每个群落的禁止捕捞和限制进入区域内至少布设了12条样带。所有横断面调查都是在上午9点到下午4点之间进行的。所有大型游动鱼类的数量和大小均在5米范围内记录,而所有小型附着于站点的礁鱼均在2米范围内记录。利用该方法将所有珊瑚鱼的长度和丰度转化为样条水平的生产力rfishprod包(Morais和Bellwood 2020),提供单个礁鱼物种的生产力估计,以及根据已公布的长度-重量关系(<一个href="https://www.fishbase.com" target="_blank" rel="noopener">https://www.fishbase.com).根据对七个SMA群落226户家庭的访问,讨论了捕鱼和鱼类消费的模式,物种也被分为目标或非目标(Parks 2017)。
然后,利用统计匹配将未采储量和PPAs内的生产力和生物量估算值与预测的反事实条件进行比较,后者代表了在缺乏管理的情况下对这些相同区域的估算值(Olmos and Govindasamy 2015, Ho et al. 2018)。我们选择了9个背景因素用于匹配,这些因素包含珊瑚礁的环境和社会特征,已知这些特征会影响响应变量或保护区的配置(附录1,表A1)。Smallhorn-West等人(2019)提供了这些变量的详细信息。这些因素包括:捕捞压力、波浪能量、深度、坡度、生境复杂性、珊瑚覆盖、生境、岛屿群和测量员。No-take横断面和PPA横断面与相同的总体对照横断面池相匹配。栖息地类型、岛屿群和测量者这些变量都是固定的,因此只有当控制样条与这些协变量的精确组合相匹配时,才能将控制样条与管理样条配对。在固定匹配之后,所有剩余的协变量被平等加权,我们通过多种匹配算法评估协变量平衡(即协变量分布在处理和控制横截面之间的差异),包括倾向评分和Mahalanobis匹配(Olmos和Govindasamy 2015)。马氏距离度量在管理和控制样条之间产生了最小的平均差异,没有证据表明无采储量或PPAs不平衡(附录1,表A1)。预先指定的容忍(即卡尺)设定了样本估计倾向得分的0.25个标准差,以确保只有高质量的匹配(Rosenbaum和Rubin 1983)。所有统计匹配程序在R 4.0.2 (R Core Team 2016)中使用MatchIt包,总共324个no-take reserve或PPA样条被匹配,只有三个剩余的不匹配(附录. 1,表A2)。
最后,利用线性混合效应模型全球语言监测机构以群落和站点为随机因子,站点嵌套在群落内,利用函数检验无采储量或PPAs与匹配的开放样条之间的目标生产力、总生产力和生物量的总体差异。模型拟合固定和随机斜率使用伽马测井链接分布和一个最低赤池信息准则(AIC)评分包括。模型拟合采用偏残差图检验,残差平方和和残差自由度采用卡方检验检验。
案例研究2:瓦努阿图的可捕性
在瓦努阿图,我们通过对鱼类的行为调查,来量化公开捕鱼区、ppa区和完全封闭区之间用鱼叉捕鱼的概率有何不同。我们建立在先前对鱼类行为的研究的基础上,这些研究测量了鱼类逃跑的距离(飞行起始距离)(Januchowski-Hartley et al. 2014),通过整合平均鱼刺距离来估计每个区域的捕获概率,并计算捕获能力随大小的变化情况。2011年12月,在瓦努阿图恩古纳-佩莱海洋保护区网络的两个社区附近进行了飞行启动距离(FID)调查(图1)。恩古纳-佩莱海洋保护区网络是一项始于2002年的协作管理倡议,包括16个村庄、政府部门、非政府组织、国际志愿者组织和研究机构。每个群落礁区都有定期收获的PPA,其中每六个月只有社区成员允许捕鱼3-7天,一个已经存在了6年的禁渔区,以及开放捕鱼的礁区(Januchowski-Hartley et al. 2014)。所有的禁捕区和保护区都将礁围在0.08到0.1平方公里之间,而公开捕鱼区则将礁围在0.14到0.16平方公里之间。我们承认,鱼叉捕鱼只是捕捞礁鱼的一种方法,但对其他方法(如线钓或网钓)的额外分析超出了本研究的范围。尽管这种抽样设计无法在统计上控制协变量,如栖息地、波浪能量或距离村庄的距离,但我们采用了一种粗匹配方法,根据专家意见,将公开捕鱼区域与管理下的珊瑚礁最相似的区域进行比较(Ahmadia等,2015)。
在每个区域,我们测量了24条鱼的FID,它们来自两个最常见的珊瑚礁鱼类类群:刺鳍鱼科(外科医生鱼和独角鱼)和红鳍鱼科(鹦嘴鱼),所有鱼的最短总长度均为15厘米(红鳍鱼科)和20厘米(红鳍鱼科)。每个目标科只对渔获物中出现的物种进行抽样,并在每个地区对相似的物种进行抽样。通过自由潜水估计FID,观察员(FAJ)从水面识别了一条焦点鱼,然后在>8米的距离下降到大约相同的深度(Januchowski-Hartley et al. 2014)。然后观察者以稳定的速度向鱼游去,直到鱼逃走。此时,观察者在他的脸正下方的基质上放置了一个标记,在鱼开始逃跑的地方放置了第二个标记,这两个点之间的距离被认为是FID。每个个体的大小(cm TL)在进入前被视觉估计。
克里尔的调查显示,珊瑚礁捕鱼的主要装备是鱼叉,在43次捕鱼中使用了33次(6次夜间鱼叉),渔网是第二大最常见的装备,在9次捕鱼中使用。Fisher的采访表明,鱼钩和鱼线在鱼叉枪航行时通常也被用作次要齿轮。在面试过程中,每位渔夫(n= 14)被要求估计鱼叉枪的最大有效射程(从渔民的脸上测量)。估计平均最大有效射程为337厘米,范围为305-360厘米。由于大多数渔民没有将鱼叉固定在鱼叉枪上,因此如果射击不成功,鱼叉就会丢失,所以在这个距离上,鱼叉成功的可能性很大。
每条鱼被分为”捕获”或”逃了出来”根据FID是否大于或小于speargun的平均、最低和最高估计范围(附录1,表A3)。首先采用z标准化的总长度和管理类型的二项logistic回归模型对所有刺鳍科和海鲂科进行了分析,以识别管理类型之间的可捕性差异,然后对不同组合的鱼类和管理类型进行了分析,以模拟鱼类大小与捕获概率之间的关系。所有分析均使用全球语言监测机构在R 4.0.2 (R核心团队2016)中发挥作用。
案例研究3:所罗门群岛的脆弱性
我们使用上述复合脆弱性指标(张等人2007年,Abesamis等人2014年)来检查所罗门群岛西部省份一个沿海社区10年共同管理期间的珊瑚礁渔业捕鱼量变化(图1)。由于社区与合作伙伴非政府组织世界鱼类组织(WorldFish)签订了保密协议,所以没有提供社区名称和位置。2008年,该社区在一个更广泛的共同管理框架内,在其传统渔场的珊瑚礁区域实施了两项周期性收获的PPAs(分别为3.6平方公里和1.4平方公里),作为非政府组织支持的计划的一部分,以制定资源使用法规和教育、合规和监测战略(Cohen et al. 2013)。在之前的研究中,这些ppa被称为禁忌或定期收获的闭包(Cohen and Alexander 2013, Cohen and Foale 2013)。自建立以来,这两个PPA区在一年中大部分时间(1月至11月)都被管理为禁止进入或使用,并定期在每年12月进行为期2周的为期1个月的收获活动,在这些活动中,捕捞既发生在PPA区礁内,也继续在邻近的礁上进行(Cohen等人,2013年)。在间歇的时间里,这些地区偶尔也会开放,为特殊的活动和宴会提供一晚上的鱼类捕捞。请注意,在开幕活动期间也经常实施额外的限制,例如禁止在保护区内用网捕鱼或夜间捕鱼。PPAs的总面积为0.86 km²,约占该群落礁渔业总面积的15%。此前对这些特定的PPAs (Cohen和Alexander 2013, Cohen和Foale 2013)和其他类似的(Cinner等人2006,Goetze等人2016,Januchowski-Hartley等人2014,Carvalho等人2019)的评估侧重于单个收获事件(Cohen和Alexander 2013, Cohen等人2013,Goetze等人2017)。在这里,我们基于这些快照研究,利用这个社区整个渔场10年期间的渔获数据,分析了渔获物组成和脆弱性的变化。
根据Cohen和Alexander(2013)以及Cohen等人(2013)的方法,从2010年到2020年,在10年的时间里,在5个采样期收集钓鱼行程和渔获数据。每个采样周期持续约1个月,与PPA开放的主要时间相对应,因此在开放事件的任何一方收集的数据至少为1周。在取样期间,我们记录了在这两个部分受保护区域(当开放时,n= 813)以及一年四季开放捕鱼的水域,n= 1369次垂钓(采样总天数为138天)由于资金和实地工作的实际情况,我们无法每年进行抽样,而且抽样间隔不均匀。在每个采样期间,至少有一名观察员日夜驻守在村庄的登陆点,一旦渔民回到岸上,他们就被要求重新描述行程的细节,包括捕鱼地点和管理区。然后使用当地语言名称记录每个捕获物的数量和物种,随后根据Cohen等人(2014年)将这些名称转换为最高的分类分辨率。然后,在Abesamis等人(2014)或张等人(2007)的补充材料中为每条鱼分配了脆弱性评分,该评分提供了最高的分类分辨率。另一项分析仅用家庭水平的分数完成,结果没有实质性的差异。
采用广义加性模型考察开放和部分保护珊瑚礁之间的平均渔获脆弱性随时间的变化,管理和时间作为固定效应。在样条内最多允许四个节的拟合,但如果能产生更好的模型拟合,则可以减少。模型采用高斯分布拟合,最终模型的选择以AIC得分最低为依据。通过检查残差直方图和拟合值与残差来评估模型拟合,这两种情况都表明模型拟合是可以接受的。所有分析均在R 4.0.2 (R CoreTeam 2016)中使用gam而且visreg功能。
结果与讨论
案例研究1:汤加的生产力
我们在汤加的调查显示,与控制区相比,禁捕保护区和ppa区都具有显著更高的渔业生产力潜力(图2a,b;附录1,表A4)。(礁鱼发展迟缓(ha)(¹day)¹±0.018 SE)在社区成员仍允许垂钓的PPAs (1.06 kg ha(¹day))内(1.20 kg ha(¹day)),与统计上匹配的允许垂钓的控制站点(0.64 kg ha(¹)(¹¹)(¹±0.076 SE))相比(0.64 kg ha((¹)(¹))发展迟缓(¹)(¹)。与此相反,与公开捕鱼区和ppa区相比,目标物种的常设生物量只在禁止捕鱼保护区观察到显著差异(图2c,d)。
经常有报告称靠近捕鱼区域内的生物量增加(MacNeil等人2015年,Cinner等人2018年,Smallhorn-West等人2020年b),但这些收益对渔业来说无关重要,除非成鱼或幼鱼补贴溢出到渔场,这方面的证据不太常见(但见Halpern等人2009,Harrison等人2012)。相比之下,在仍有一些捕鱼活动的这些ppa区域,生产力的提高应会导致渔民的渔获量的改善,即使常设生物量仍然低于全面禁止捕鱼所能达到的水平。本研究所研究的PPAs与开放区域的目标物种生产力差异为0.42 kg ha(¹day)¹,即生产力增加65%,这可能表明部分保护可能增加抽取率。当然,如果PPA礁鱼类群落恢复持续,鱼类群落的大小结构也会向更大的鱼类转变,这些鱼类对潜在日增长的贡献较小(Morais和Bellwood 2020)。然而,考虑到汤加的社区网络强制准入限制,现在总共覆盖了96.9平方公里的珊瑚礁面积(Smallhorn-West等人,2020年一个),我们可以合理地预期,如果这片珊瑚礁区域在联合管理下在全国范围内开放捕鱼,那么每天的鱼类产量将会高出约4.1吨。
案例研究2:瓦努阿图的可捕性
我们的调查显示,在不同的管理策略之间,棘刺科和海蟹科的捕获能力存在大小依赖性(图3;附录1,表A5)。随着捕鱼限制的增加(从公开捕捞的珊瑚礁到PPA,再到禁止捕捞的保护区),个体接近鱼叉范围的概率也在增加(附录1,表A5)。例如,对于22厘米(TL)的(在本研究刺尾鱼科的平均大小),捕获范围内接近的概率为0.22(0.12 - -0.39)在开放的珊瑚礁,在PPA 0.56(0.40 - -0.70),和0.85(0.72 - -0.93 95%可信区间)禁捕区(图3)。同样,鹦嘴鱼的平均大小(26厘米TL),在开放的珊瑚礁,据预测,1在10(成功概率0.11,0.04 - -0.26)方法会成功,而在PPA率为1 2 (0.48,0.34-0.63范围),而对于不接收的储量,10个接近(0.82,0.74-0.89范围)中有8个是成功的。考虑到标枪距离的上下范围(分别为3.6 m和3.05m)对总体趋势没有影响(附图1,图2)。A1, A2)。
可捕性的差异也是鱼大小的产物,较大的鱼通常更难以接近。然而,随着捕鱼限制的增加,这种模式发生了变化,在保护区和禁捕保护区中,中等大小的鱼类往往不如在开放捕鱼区抽样的最小的鱼类警惕。相反,较小的个体通常在所有管理形式中都不那么谨慎。在公开捕捞和部分保护的珊瑚礁上,由于捕鱼的选择性压力,更大、更古老的鱼更容易警惕和难以接近,容易接近的鱼可能在完全成熟之前就被杀死了。不同管理区之间捕获量大小的差异可能是由鱼类个体行为的变化所驱动的,而不是由渔业驱动的选择所驱动的,因为在短期捕鱼事件后,增加的警惕是显而易见的,这些事件只会清除一小部分正常种群,并且naïveté在关闭时间短于个体生命周期后返回(Januchowski-Hartley et al. 2014)。
在各种形式的共同管理下,捕获量的变化对渔业有不同的短期和长期影响。重要的是,这些周期性采收的PPAs的关闭时间为6个月,远远短于棘叶目或疤叶目种群恢复所需的时间(Abesamis等人2014,Januchowski-Hartley等人2014)。因此,虽然在短期内,由于鱼类naïveté而提高了这些珊瑚礁的捕获力可能会给渔民带来巨大的利益,但这可能会损害渔业的长期可持续性,以及生态系统的整体状态,这取决于捕捞总量。这种行为变化可能会产生一种形式的超稳定性,即高产掩盖了耗尽的丰度,造成渔业状况比实际状况更好的错误印象(Erisman等人2011年,Hamilton等人2016年,Maggs等人2016年)。相反,捕鱼量的变化也可能会导致公开捕捞区域的过度枯竭,因此,由可怕的鱼类导致的低产量意味着比实际低的储量状态(Lennox et al. 2017)。因此,为了妥善管理这些权衡,PPA开口的频率和持续时间需要进一步考虑(Goetze等人2016、2017、2018,Carvalho等人2019)。通常情况下,PPA开口是对社区需求或愿望的回应,但将它们与目标物种的生活史特征相匹配更有可能提高其使用的可持续性(Cohen和Foale, 2013年)。
案例研究3:所罗门群岛的脆弱性
对五个最常见捕捞鱼类科的分析显示,PPAs渔获量的组成发生了重大变化(图4a)。在10年的研究期间,在定期捕捞的珊瑚礁上,最常见的鱼类family Acanthuridae(占10年期间总捕捞量的31%)的比例从54%下降到3%。同时,epinephelid的比例(占总渔获量的9%)从4%增加到34%。额外的分析表明,这些变化也与捕获的绝对个体数量一致,在PPAs中,每次捕获的刺鳍鱼平均数量从8.5(±1.1 SE)下降到0.26(±0.26 SE),而在PPAs中捕获的epinephelids平均数量从0.77(±0.12 SE)上升到2.42(±1.2 SE)(附录1,图A3)。这些变化在一年四季持续开放捕鱼的珊瑚礁上并不明显。随着时间的推移,渔获物组成的变化也改变了渔获物的整体生活史特征。到2019年(PPA实施10年后),登陆渔获量比2010年增加了30%(即,它们构成了更大比例的具有脆弱生活史特征的物种),但对开放持续捕鱼的珊瑚礁渔获量保持相对稳定(图4b;附录1,表A6)。脆弱物种的增加表明,渔民现在从PPA捕获的物种比例比以前更多,这些物种具有生活史特征,容易被过度捕捞(即那些生长较慢或初次成熟较老的物种)。这是由捕获的低脆弱性刺叶目相对丰度的下降(平均脆弱性26.9)和高脆弱性epinephelid(平均脆弱性55.2)的相对丰度的增加所驱动的。 The increase in epinephelid catch was also largely driven by benthic species of the genusCephalopholis(平均漏洞46.0)和Epinephelus脆弱性(平均63.0)。值得注意的是,专注于单个开始/收获事件而不是时间序列的研究得出的结论是,周期性收获的PPAs可以支持较低脆弱性物种相对较高的生物量(Jupiter等人2012,Januchowski-Hartley等人2014,Goetze等人2016)。另外值得注意的是,尽管进行了10年的积极管理,但其他常见鱼类家族的渔获量并没有增加。
渔获量数据是对一个区域内种群特征的一种依赖渔业的衡量,因此,一系列因素可能会混淆渔获量与种群状况之间的关系(Maggs等人,2016)。这些观察到的模式可能是由于(1)高脆弱性物种的增加,(2)低脆弱性物种的减少,或(3)渔民或鱼类行为的改变。epinephelids(高脆弱性)和acanthurids(低脆弱性)捕鱼量的变化可能是由于物种丰度的变化,要么通过种群恢复/损失,要么通过溢出(个体聚集在压力较小的区域)(Halpern et al. 2009)。以较小的底栖epinephelid物种为目标也表明,在监测开始之前,较大的石斑鱼可能已经灭绝,因为它们通常是珊瑚礁上的第一个目标物种(Coleman et al. 2000, de Mitcheson et al. 2020)。如果是这样的话,那么我们的发现是先前最大、最脆弱的物种灭绝的产物,接着是大量低脆弱性物种的优先选择和灭绝,然后是进一步转向更脆弱、但以前未被充分利用的群体。或者,可能是渔民的行为发生了变化(例如,渔具使用的变化或对不同鱼种的感知价值的变化)或鱼的行为发生了变化(例如,可捕性的变化)。例如,渔民可以主动瞄准PPAs内的棘螈,或被动瞄准这些物种,因为它们在PPAs内行为的变化使它们相对容易被捕获。在我们的研究设计中使用渔获数据的局限性使我们无法确定哪一种解释是最正确的。在这个社区中,Cohen等人(2013)也证明了开放后两周内PPA区域内的捕鱼压力可能比临近连续捕鱼的珊瑚礁全年的捕鱼压力更大。
无论涉及的确切机制是什么,如果管理的目标是可持续地使用一种资源,那么,渔获物生活史特征向更脆弱和寿命更长的物种的变化意味着需要对现有规则进行适应性管理。在这种情况下,调整管理规则将需要调整PHC开口的努力、周期性和持续时间与目标物种的生活史特征。这有两种可能发生的方式:首先,如果偏好特定的物种,那么可以调整收获的时期,以便与这些物种的生活史保持一致。或者,如果社会文化规则限制采收发生的时间,那么将采收限制在在该周期下可持续的特定分类群将是一个可行的替代方案。无论哪种选择,这都强调共同管理的适应性,包括增加关闭期的持续时间,减少开放期的持续时间,和/或对捕鱼努力量和收成进一步增加投入或产出控制。这些反馈机制使管理部门能够随着时间的推移适应不断变化的环境和种群特征,最终将影响渔民是否在长期内获得相对稳定的渔获量和渔获率(Armitage et al. 2008)。
结论
通过对三个太平洋岛国近3000项观察结果的分析,我们揭示了部分保护可以影响自然资源的使用和可持续性的三种方式(生产率、可获取性和脆弱性)。在汤加的案例研究中,我们发现PPAs内的潜在渔业生产力更高,这表明渔民的捕获量得到了提高。在瓦努阿图的案例研究中,执行公私合作协定使单条鱼更容易用矛捕获,这应使渔民获得更大的捕鱼效率。然而,如果对努力量或渔获量没有额外的控制,增加的可捕性也会使目标物种更容易过度捕捞。在所罗门群岛的案例研究中,渔获物的构成在十多年来改变了部分保护的管理方式,并由本质上更容易受到过度开发的物种主导。每一个小故事都说明了可以通过对资源及其用户之间的反馈和学习做出响应的适应性共同管理的不同方面来管理的不同的好处和风险(Armitage等人2008)。
我们发现,尽管部分保护可以给渔民和渔业带来明显的好处,例如提高生产力和捕获量,但这种管理方法也可能放大某些风险。PPAs中鱼类行为的大小依赖变化可能会诱发一种形式的超稳定性,在这种情况下,产量和丰度变得脱钩,即使种群下降,渔获量也可能保持稳定(Erisman等人2011年,Hamilton等人2016年,Maggs等人2016年)。过度稳定通常与聚集的物种有关,比如北大西洋鳕鱼,这导致了渔业的崩溃(罗斯和库尔卡,1999)。如果PPA区内鱼类行为的变化速度快于种群恢复速度,PPA区内较大的个体比其边界外的较小的个体更容易捕获,则PPA区内大型鱼类的捕获量可能反映的是驯服性的增加而不是丰度的增加。在渔获样本不能反映真实种群状况的情况下,这可能会造成对增长和种群恢复的错误认识。其次,在十年的时间尺度上,ppa内脆弱物种捕鱼量的比例增长本身就表明了一种潜在的风险,因为管理的最初目标可能没有计划应对这些变化。
PPAs的适应性共同管理是一种机制,通过它可以减轻相关的风险——过度稳定和过度开发脆弱物种——同时获得部分保护的渔业利益。管理的适应性本质意味着一个基于新的经验和见解(如监测)的不断变化的管理实践的迭代过程,以引导管理系统走向期望的状态(Cinner等,2019)。在我们的案例中,与ppa相关的风险适应性管理包括调整关闭和打开的周期(例如,从每6个月一次到每年一次,或从每年一次到每2年一次),以及开发的强度或方法(例如,袋或尺寸限制,或对特定捕鱼方法的限制),在我们的案例研究中,这些变化通常是基于社会需求和需求,而不是基于所使用资源的可持续性(Cohen和Steenbergen, 2015)。不管具体的机制是什么,PPAs的长期可持续性可能取决于反馈机制,使管理层能够适应不断变化的环境和股票特征(Armitage et al. 2008)。
虽然我们对ppa的好处和风险的调查引起了人们对资源使用者将如何体验以前被忽视的影响模式的关注,但未来的研究仍有几个机会通过不同的研究设计来扩展这些见解。首先,在沿海社区和政府看来,我们审查的所有PPAs都是已建立的共同管理系统的一部分,具有很高的合法性(Januchowski-Hartley等人2014年,Cohen和Steenbergen等人2015年,Smallhorn-West等人2020年一个).如果PPAs是通过自上而下的治理机制指定、实施和强制执行的,在这种情况下,人们认为规则的创建不公平(Mascia et al. 2010),或者在政府眼中合法性较低的本地管理系统中,或者管理配置出于政治动机,以尽量减少与资源使用的重叠(Cockerell et al. 2020),那么结果可能会不同。其次,这项研究的数据局限性意味着我们不能采用完全交叉抽样设计,这将使我们能够在部分保护下评估驱动渔获物组成和种群动态随时间变化的精确机制。例如,在所罗门群岛的案例研究中,同时使用紫外线和行为数据将使我们能够确定渔获成分的时间趋势是由于鱼类或渔民行为的变化,还是生态系统的变化。利用10年时间尺度上同一地点与渔业相关和独立的数据来研究这些新指标如何相互作用,将大大提高我们对海洋管理影响途径的理解。我们也承认,案例研究中的观察结果往往具有很强的情境依赖性,产生可推广的知识主张需要从个体案例研究中扩展观察结果和推论,超越最初产生案例的空间和/或时间边界(Magliocca et al. 2018)。此外,我们在这里关注的是PPAs的渔业结果,但其他社会经济影响,以及与生态完整性、功能和保护结果相关的影响,也都是至关重要的理解(Cinner et al. 2012)。最后,量化挖角和合规的程度,以及它们对我们研究结果的潜在影响,超出了本研究的范围。
最后,这项研究增加了保护区作为自然资源管理工具的细微差别和复杂性。我们证明,在某些情况下,部分保护能够改变渔业的表现——通过改变生态系统动态、鱼的行为方式和人们捕获的东西。但我们也承认,这些变化的净效应好坏参半,除了渔业指标之外,还需要进一步考虑。即使有积极的渔业利益,如果这些渔业利益是以自然为代价的,则管理海洋系统中的其他利益相关者也可能受到不利影响。保护问题也继续存在,因此完全封闭的禁止捕捞管理保护区必须继续成为空间管理的组成部分,因为它们可以提供积极的保护和渔业影响(例如,Harrison等人2012年,Edgar等人2014年)。最后,全球管理区域中只有一小部分(> %)是通过自下而上或共同管理机构完成的(Gurney等人,2021年),目前尚不清楚这些指标中的一部分或全部是否会受到自上而下治理框架的类似影响。这些小故事表明,在有效管理的情况下,共同管理或面向社区的公私合作协定可以为渔业提供具体的好处,但前提是要清楚地了解潜在的风险。
对本文的回应
欢迎对本文作出回应。如果被接受发表,您的回复将被超链接到文章。<一个href="//www.dpl-cld.com/responses.php?articleid=13112&mode=add">要提交响应,请点击此链接.<一个href="//www.dpl-cld.com/responses.php?articleid=13112">要阅读已经接受的回复,请点击此链接.致谢
这项研究由澳大利亚研究委员会、国家地理学会和澳大利亚政府通过澳大利亚国际农业研究中心(ACIAR)项目FIS/2016/300资助。这项工作是作为世界鱼类协会领导的国际农业研究磋商小组(CGIAR)鱼类农业粮食系统研究项目(Fish)的一部分进行的。该项目得到了CGIAR信托基金捐助方的支持。
数据可用性
稿件通过审核后,数据和代码将被上传至在线数据库PANGAEA
文献引用
阿贝萨米斯,R. A., A. L.格林,G. R. Russ, C. R. L. Jadloc. 2014。珊瑚礁鱼类对捕捞的内在脆弱性及其在渔业关闭中恢复的差异。海洋生物学报24(4):1033- 1033。<一个href="https://doi.org/10.1007/s11160-014-9362-x" target="_blank" rel="noopener">https://doi.org/10.1007/s11160-014-9362-x
Ahmadia, G. N., L. Glew, M. Provost, D. Gill, N. I. Hidayat, S. Mangubhai, Purwanto和H. E. Fox. 2015。将影响评估纳入海洋保护区监测的设计和实施。英国皇家学会哲学学报B:生物科学370::20140275。<一个href="https://doi.org/10.1098/rstb.2014.0275" target="_blank" rel="noopener">https://doi.org/10.1098/rstb.2014.0275
Armitage, D., M. Marschke和R. Plummer, 2008。适应性共同管理与学习的悖论。全球环境变化18(1):86-98。<一个href="https://doi.org/10.1016/j.gloenvcha.2007.07.002" target="_blank" rel="noopener">https://doi.org/10.1016/j.gloenvcha.2007.07.002
Ban, n.c., G. G. Gurney, n.a. Marshall, C. K. Whitney, M. Mills, S. Gelcich, n.j. Bennett, M. C. Meehan, C. Butler, S. Ban, T. C. Tran, M. E. Cox和S. J. Breslow。2019。海洋保护区的福祉结果。自然的可持续性2:524 - 532。<一个href="https://doi.org/10.1038/s41893-019-0306-2" target="_blank" rel="noopener">https://doi.org/10.1038/s41893-019-0306-2
布洛克,r.e., 1985年。夏威夷鱼类聚集装置附近远洋鱼类摄食习性的初步研究,或鱼类聚集装置能否提高本地渔业生产力?海洋通报37(1):40-49。
Brockington, D.和K. Schmidt-Soltau。2017.荒野和发展对社会和环境的影响。羚羊38(2):140 - 142。<一个href="https://doi.org/10.1017/S0030605304000250" target="_blank" rel="noopener">https://doi.org/10.1017/S0030605304000250
卡瓦略,P. G., J. Goetze, S. D. Jupiter, J. Claudet, F. A. J. Hartley, R. Weeks. 2019。通过定期关闭渔场优化捕捞。应用生态学报56(8):1927-1936。<一个href="https://doi.org/10.1111/1365-2664.13417" target="_blank" rel="noopener">https://doi.org/10.1111/1365-2664.13417
张,W. W. L., T. J.皮彻,D.保利,2005。海洋鱼类渔业固有灭绝脆弱性评估的模糊逻辑专家系统。生物保护124(1):97 - 111。<一个href="https://doi.org/10.1016/j.biocon.2005.01.017" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2005.01.017
张,W. W. L., R. Watson, T. Morato, T. J. Pitcher和D. Pauly. 2007。全球捕鱼量的内在脆弱性。海洋生态进展系列333:1-12。<一个href="https://doi.org/10.3354/meps333001" target="_blank" rel="noopener">https://doi.org/10.3354/meps333001
Cinner, j.e., j.d. Lau, a.g. Bauman, d.a. Feary, f.a. Januchowski-hartley, c.a. Rojas和m.l. Barnes. 2019。16年的社会和生态动态揭示了在维持公地方面适应性管理的挑战和机遇。美国国家科学院学报116(52):26474-26483。<一个href="https://doi.org/10.1073/pnas.1914812116" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.1914812116
辛纳、J. E.、E.梅尔、C.胡奇瑞、M. A.麦克尼尔、N. A. J.格雷厄姆、C.莫拉、T. R.麦克拉纳汉、M. L.巴恩斯、J. N.基廷格和C. C.希克斯。人类影响的严重性调节了珊瑚礁保护的成效。美国国家科学院学报115(27):E6116-E6125。<一个href="https://doi.org/10.1073/pnas.1708001115" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.1708001115
Cinner, J., M. J. Marnane, T. R. McClanahan和G. R. Almany, 2006。印度洋-太平洋地区适应性珊瑚礁管理的阶段性关闭。生态与社会11(1):31。<一个href="https://doi.org/10.5751/ES-01618-110131" target="_blank" rel="noopener">https://doi.org/10.5751/ES-01618-110131
辛纳,J. E., T. R. Mcclanahan, M. A. Macneil, N. A. J. Graham, T. M. Daw, A. Mukminin, D. A. Feary, A. L. Rabearisoa, A. Wamukota, N. Jiddawi和S. J. Campbell。珊瑚礁社会-生态系统的管理。美国国家科学院学报109(14):5219-5222。<一个href="https://doi.org/10.1073/pnas.1121215109" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.1121215109
cin, j . E。j . Zamborain-Mason g·g·格尼,n . a·j·格雷厄姆·m·a·麦克尼尔a·s·霍利,c·莫拉农村,E . Maire t·r·麦克拉纳罕j·m·麦纳j . n .基廷格c·c·希克斯,s D 'Agata c . Huchery m·l·巴恩斯D . a . Feary i D·威廉姆斯,m . Kulbicki l . Vigliola l . Wantiez g·j·埃德加·r·D·stuart smith, a . Sandin, a . l .绿色m .乞讨,a . m .弗里德兰德,美国k·威尔逊,E . Brokovich a·j·布鲁克斯,j . j . Cruz-Motta D·j·布斯,p . Chabanet m . m s c a . FerseU. R. Sumaila, M. J. Hardt和D. Mouillot, 2020。在人类主导的世界中实现渔业、生态系统功能和生物多样性目标。科学》368(6488):307 - 311。<一个href="https://doi.org/10.1126/science.aax9412" target="_blank" rel="noopener">https://doi.org/10.1126/science.aax9412
cokerell, B., R. L. Pressey, A. Grech, J. G. Álvarez-Romero, T. Ward, R. Devillers, 2020。代表权并不一定会减少对生物多样性的威胁:澳大利亚联邦海洋保护区系统,2012-2018年。生物保护252:108813。<一个href="https://doi.org/10.1016/j.biocon.2020.108813" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2020.108813
科恩(P. J. Cohen)和亚历山大(T. J. Alexander), 2013年。通过定期关闭渔场来管理珊瑚礁的渔获率、组成和鱼的大小。PLoS ONE 8(9): e73383。<一个href="https://doi.org/10.1371/journal.pone.0073383" target="_blank" rel="noopener">https://doi.org/10.1371/journal.pone.0073383
Cohen, P. J., J. E. Cinner, S. Foale, 2013。与海洋休渔期相关的渔业动态。全球环境变化23(6):1702-1713。<一个href="https://doi.org/10.1016/j.gloenvcha.2013.08.010" target="_blank" rel="noopener">https://doi.org/10.1016/j.gloenvcha.2013.08.010
Cohen, P. J.和S. J. Foale, 2013。在定期捕捞的海洋保护区维持小规模渔业。海洋政策37:278 - 287。<一个href="https://doi.org/10.1016/j.marpol.2012.05.010" target="_blank" rel="noopener">https://doi.org/10.1016/j.marpol.2012.05.010
Cohen, P. J.和D. J. Steenbergen. 2015。珊瑚三角地区渔业共同管理的社会层面。环境保护42(3):278 - 288。<一个href="https://doi.org/10.1017/S0376892914000423" target="_blank" rel="noopener">https://doi.org/10.1017/S0376892914000423
Cohen, P., S. Tapala, A. Rikio, E. Kukiti和F. Sori。2014。建立对所罗门群岛维拉拉维拉北部渔业管理分类学的共同认识。传统海洋资源管理与知识信息通报33(6):3-12。
科尔曼,F. C., C. C.凯尼格,G. R.洪博培,J. A. Musick, A. M. Eklund, J. C. McGovern, G. R. Sedberry, R. W. Chapman和C. B. Grimes。2000。长寿的礁鱼:石斑鱼-鲷鱼群。25(3):渔业的程度。<一个href="https://doi.org/10.1577/1548-8446(2000)025<0014:LRF>2.0.CO;2" target="_blank" rel="noopener">https://doi.org/10.1577/1548 - 8446(2000) 025 < 0014:探测器> 2.0.CO; 2
de Mitcheson, Y. J. S., C. Linardich, J. P. Barreiros, G. M. Ralph, A. Aguilar-Perera, P. Afonso, B. E. Erisman, D. A. Pollard, S. T. fenessy, A. A. Bertoncini。有价值但又脆弱:过度捕捞和管理不足继续威胁着石斑鱼,那么现在该怎么办?海洋政策116:103909。<一个href="https://doi.org/10.1016/j.marpol.2020.103909" target="_blank" rel="noopener">https://doi.org/10.1016/j.marpol.2020.103909
埃德加,G. J., R. D.斯图尔特-史密斯,T. J.威利斯,S. Kininmonth, S. C.贝克,S. Banks, n.s. Barrett, M. A. Becerro, A. T. F. Bernard和J. Berkhout. 2014。全球保护成果取决于具有五个关键特征的海洋保护区。自然506(7487):216 - 220。<一个href="https://doi.org/10.1038/nature13022" target="_blank" rel="noopener">https://doi.org/10.1038/nature13022
埃里斯曼,b.e., L. G. Allen, J. T. Claisse, D. J. Pondella, E. F. Miller, J. H. Murray。富足的假象:过度稳定掩盖了两个以鱼类产卵群为目标的休闲渔业的崩溃。中国水产科学学报,29(5):393 - 393。<一个href="https://doi.org/10.1139/f2011-090" target="_blank" rel="noopener">https://doi.org/10.1139/f2011-090
费拉罗,P. J., M. M.哈诺尔,2011。通过公园和保护区保护生态系统,减轻贫困:”双赢”还是权衡?环境与资源经济学48:269-286。<一个href="https://doi.org/10.1007/s10640-010-9408-z" target="_blank" rel="noopener">https://doi.org/10.1007/s10640-010-9408-z
芬尼,b.p., I. Gregory-Eaves, M. S. V.道格拉斯,J. P. Smol, 2002。在过去的2200年里东北太平洋的渔业生产力。自然416(6882):729 - 733。<一个href="https://doi.org/10.1038/416729a" target="_blank" rel="noopener">https://doi.org/10.1038/416729a
Foale, S., P. Cohen, S. Januchowski-Hartley, A. Wenger, M. Macintyre。2011。保留区和禁忌:太平洋渔业的起源和影响。渔业12(4):357-369。<一个href="https://doi.org/10.1111/j.1467-2979.2010.00395.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1467-2979.2010.00395.x
Giakoumi, S., C. Scianna, J. Plass-Johnson, F. Micheli, K. Grorud-Colvert, P. Thiriet, J. Claudet, G. Di Carlo, A. Di Franco, S. D. Gaines, J. A. García-Charton, J. Lubchenco, J. Reimer, E. Sala, P. Guidetti. 2017。在拥挤的地中海全面和部分保护的生态影响:区域荟萃分析。科学报告7(1):1 - 12。<一个href="https://doi.org/10.1038/s41598-017-08850-w" target="_blank" rel="noopener">https://doi.org/10.1038/s41598-017-08850-w
格策、J. S.、J. Claudet、F. Januchowski-Hartley、T. J. Langlois、S. K. Wilson、C. White、R. Weeks和S. D. Jupiter。2018。展示定期关闭渔场的多重好处。应用生态学报55(3):1102-1113。<一个href="https://doi.org/10.1111/1365-2664.13047" target="_blank" rel="noopener">https://doi.org/10.1111/1365-2664.13047
Goetze, j.s., f.a. Januchowski-Hartley, J. Claudet, T. J. Langlois, S. K. Wilson, S. D. Jupiter. 2017。鱼类警惕度是一个比丰度、长度或生物量变化更敏感的指标。生态学应用27(4):1178 - 1189。<一个href="https://doi.org/10.1002/eap.1511" target="_blank" rel="noopener">https://doi.org/10.1002/eap.1511
Goetze, J., T. Langlois, J. Claudet, F. Januchowski-Hartley, S. D. Jupiter. 2016。定期封育需要对脆弱物种进行充分保护,并延长封育期。生物保护203:67 - 74。<一个href="https://doi.org/10.1016/j.biocon.2016.08.038" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2016.08.038
格尼、G. G. E. S.达林、G. N.艾哈迈德迪亚、V. N.阿戈斯蒂尼、N. C.班、J.布莱思、J.克劳德特、G.爱普斯坦、埃斯特拉迪瓦里、A.希姆斯-康奈尔、H. D.乔纳斯、D.阿米蒂奇、S. J.坎贝尔、C.考克斯、W. R.弗里德曼、D.吉尔、P.莱斯塔里、S.曼古布海、E.麦克劳德、N. A.穆蒂加、J.纳格阿、R.拉纳沃森、A.温、I.尤利安托和S. D.朱庇特。生物多样性需要所有现成的工具:利用生物化学物质。自然595(7869):646 - 649。<一个href="https://doi.org/10.1038/d41586-021-02041-4" target="_blank" rel="noopener">https://doi.org/10.1038/d41586-021-02041-4
Haasnoot, M., R. Biesbroek, J. Lawrence, V. Muccione, R. Lempert和B. Glavovic, 2020。确定加速适应气候变化的解决空间。beplay竞技区域环境变化20(2):1-5。<一个href="https://doi.org/10.1007/s10113-020-01623-8" target="_blank" rel="noopener">https://doi.org/10.1007/s10113-020-01623-8
Halpern, b.s., C. J. Klein, C. J. Brown, M. Beger, H. S. Grantham, S. Mangubhai, M. Ruckelshaus, V. J. Tulloch, M. Watts和C. White。2013。在面对社会公平、经济回报和环境保护的内在权衡时,实现三重底线。美国国家科学院学报110(15):6229-6234。<一个href="https://doi.org/10.1073/pnas.1217689110" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.1217689110
Halpern, b.s., s.e. Lester和j.b. Kellner, 2009。海洋保护区的溢出和渔业资源的补充。环境保护36(4):268 - 276。<一个href="https://doi.org/10.1017/S0376892910000032" target="_blank" rel="noopener">https://doi.org/10.1017/S0376892910000032
Hamilton, R. J., G. R. Almany, D. Stevens, M. Bode, J. Pita, N. A. Peterson, J. H. Choat. 2016。高稳定性掩盖了隆头鹦嘴鱼的数量下降(Bolbometopon muricatum)的人口。珊瑚礁35(3):751 - 763。<一个href="https://doi.org/10.1007/s00338-016-1441-0" target="_blank" rel="noopener">https://doi.org/10.1007/s00338-016-1441-0
Harrison, H. B., D. H. Williamson, R. D. Evans, G. R. Almany, S. R. Thorrold, G. R. Russ, K. A. Feldheim, L. van Herwerden, S. Planes, M. Srinivasan. 2012。海洋保护区的幼鱼出口及鱼类和渔业的补充效益。当代生物学22(11):1023 - 1028。<一个href="https://doi.org/10.1016/j.cub.2012.04.008" target="_blank" rel="noopener">https://doi.org/10.1016/j.cub.2012.04.008
Ho, D., K. Imai, G. King, E. Stuart和A. Whitworth, 2018。MatchIt:参数因果推理描述的非参数预处理。统计软件学报42(8):1-28。<一个href="https://doi.org/10.18637/jss.v042.i08" target="_blank" rel="noopener">https://doi.org/10.18637/jss.v042.i08
Januchowski-Hartley, f.a., j.e. Cinner, n.a. J. Graham. 2014。渔业受益于鱼类行为的改变,定期关闭渔场。保护水生24(6):790:777 - 790。<一个href="https://doi.org/10.1002/aqc.2388" target="_blank" rel="noopener">https://doi.org/10.1002/aqc.2388
Januchowski-Hartley, f.a., n.a. J. Graham, J. E. Cinner, G. R. Russ。2013。鱼类溢出naïveté从海洋保护区。生态学通讯16(2):191 - 197。<一个href="https://doi.org/10.1111/ele.12028" target="_blank" rel="noopener">https://doi.org/10.1111/ele.12028
Januchowski-Hartley, f.a., n.a. J. Graham, J. E. Cinner和G. R. Russ。2015。当地的捕鱼活动影响了印度洋-太平洋保护区内的珊瑚礁鱼类行为。生物保护182:8-12。<一个href="https://doi.org/10.1016/j.biocon.2014.11.024" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2014.11.024
Januchowski-Hartley, f.a., K. L. Nash, R. J. Lawton, 2012。鱼枪、潜水装备和观测者对估计鱼在珊瑚礁上飞行起始距离的影响。海洋生态进展系列469:113-119。<一个href="https://doi.org/10.3354/meps09971" target="_blank" rel="noopener">https://doi.org/10.3354/meps09971
Jennings, S.和N. V. C. Polunin, 1996。捕捞努力度和渔获率对斐济礁鱼群落结构和生物量的影响应用生态学报:400-412。<一个href="https://doi.org/10.2307/2404761" target="_blank" rel="noopener">https://doi.org/10.2307/2404761
Joppa, L. N.和A. Pfaff, 2011。对全球保护区的影响。英国皇家学会学报B辑:生物科学278(1712):1633-1638。<一个href="https://doi.org/10.1098/rspb.2010.1713" target="_blank" rel="noopener">https://doi.org/10.1098/rspb.2010.1713
朱庇特,S. D., P. J. Cohen, R. Weeks, A. Tawake, H. Govan. 2014。地方管理的海洋区域:多种目标和不同的战略。海洋生物学报20(2):329 - 331。<一个href="https://doi.org/10.1071/PC140165" target="_blank" rel="noopener">https://doi.org/10.1071/PC140165
朱庇特,S. D., R. Weeks, A. P. Jenkins, D. P. Egli, A. Cakacaka, 2012。一次集中捕捞事件对传统海洋封闭内鱼类种群的影响。珊瑚礁31(2):321 - 334。<一个href="https://doi.org/10.1007/s00338-012-0888-x" target="_blank" rel="noopener">https://doi.org/10.1007/s00338-012-0888-x
Lennox, R. J., J. Alós, R. Arlinghaus, A. Horodysky, T. Klefoth, C. T. Monk和S. J. Cooke. 2017。是什么让鱼容易被鱼钩捕获?一个概念框架和对关键决定因素的审查。渔业与水产18(5):986-1010。<一个href="https://doi.org/10.1111/faf.12219" target="_blank" rel="noopener">https://doi.org/10.1111/faf.12219
莱斯特,s。E。和b。s。哈尔彭,2008。海洋禁捕保护区与部分保护区的生物响应海洋生态进展系列367:49-56。<一个href="https://doi.org/10.3354/meps07599" target="_blank" rel="noopener">https://doi.org/10.3354/meps07599
麦克尼尔,m.a., N. A. J. Graham, J. E. Cinner, S. K. Wilson, I. D. Williams, J. Maina, S. Newman, A. M. Friedlander, S. Jupiter, N. V. C. Polunin。2015。世界珊瑚礁鱼类的恢复潜力。自然520(7547):341 - 344。<一个href="https://doi.org/10.1038/nature14358" target="_blank" rel="noopener">https://doi.org/10.1038/nature14358
马格斯,J. Q., B. Q. Mann, W. M. Potts, S. W. Dunlop. 2016。传统的管理策略无法阻止一种具有高度稳定性的标志性海洋休闲渔业物种的单位努力渔获量下降。渔业管理和生态23(3-4):187-199。<一个href="https://doi.org/10.1111/fme.12125" target="_blank" rel="noopener">https://doi.org/10.1111/fme.12125
Magliocca, n.r., E. C. Ellis, G. R. Allington, A. de Bremond, J. DellAngelo, O. Mertz, P. Messerli, P. Meyfroidt, R. Seppelt和P. H. Verburg。缩小全球知识差距:从社会-生态系统的案例研究中产生广义的知识。全球环境变化50:1-14。<一个href="https://doi.org/10.1016/j.gloenvcha.2018.03.003" target="_blank" rel="noopener">https://doi.org/10.1016/j.gloenvcha.2018.03.003
Mascia, m.b., c.a. Claus, R. Naidoo, 2010。海洋保护区对渔业社区的影响。保护生物学24(5):1424 - 1429。<一个href="https://doi.org/10.1111/j.1523-1739.2010.01523.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1523-1739.2010.01523.x
McClanahan, T. R., c.a. Abunge和J. E. Cinner, 2012。肯尼亚渔民和管理者对管理限制和利益偏好的异质性。环境保护39(4):357 - 369。<一个href="https://doi.org/10.1017/S0376892912000197" target="_blank" rel="noopener">https://doi.org/10.1017/S0376892912000197
Morais, r.a.和d.r. Bellwood, 2019年。在退化的珊瑚礁上,海洋补贴支撑着鱼类的生产力。当代生物学29(9):1521 - 1527。<一个href="https://doi.org/10.1016/j.cub.2019.03.044" target="_blank" rel="noopener">https://doi.org/10.1016/j.cub.2019.03.044
Morais, r.a.和d.r. Bellwood, 2020年。估计珊瑚礁鱼类生产力的原则。珊瑚礁39(5):1221 - 1231。<一个href="https://doi.org/10.1007/s00338-020-01969-9" target="_blank" rel="noopener">https://doi.org/10.1007/s00338-020-01969-9
Morais, r.a., s.r. Connolly和d.r. Bellwood, 2020年。人类的开发塑造了珊瑚礁上生产力与生物量的关系。全球变化与生物学报26(3):1295- 1295。<一个href="https://doi.org/10.1111/gcb.14941" target="_blank" rel="noopener">https://doi.org/10.1111/gcb.14941
Morais, r.a., a.c. Siqueira, p.f. Smallhorn-West和d.r. Bellwood, 2021年。空间补贴推动了热带海洋生物量生产的“甜蜜点”。PLoS Biology 19(11): e3001435。<一个href="https://doi.org/10.1371/journal.pbio.3001435" target="_blank" rel="noopener">https://doi.org/10.1371/journal.pbio.3001435
奥尔莫斯,A.和P. Govindasamy, 2015。倾向评分:一个实用的介绍使用R.多学科评价杂志11(25):68-88。
公园,j . 2017。作为汤加气候恢复部门组成部分的瓦乌特别管理区的基线社会经济调查。亚洲开发银行报告,Mandaluyong,大马尼拉,菲律宾。
R核心团队。2016。R:统计计算的语言和环境。R统计计算基础。2017。奥地利的维也纳。
Robinson, J., n.a. J. Graham, J. E. Cinner, G. R. Almany, P. Waldie. 2015。鱼类及渔民的行为影响石斑鱼的易损性(Epinephelidae)到多物种产卵聚集地捕鱼。珊瑚礁34(2):371 - 382。<一个href="https://doi.org/10.1007/s00338-014-1243-1" target="_blank" rel="noopener">https://doi.org/10.1007/s00338-014-1243-1
罗斯,g.a.和d.w.库尔卡,1999年。鱼类和渔业的超聚集:单位努力捕鱼量如何随着北方鳕鱼(Gadus morhua)拒绝了。中国水产科学,29(1):62 - 63。<一个href="https://doi.org/10.1139/f99-207" target="_blank" rel="noopener">https://doi.org/10.1139/f99-207
罗森鲍姆和d·b·鲁宾1983年著。在因果效应的观察性研究中倾向评分的中心作用。70(1):生物统计学41-55。<一个href="https://doi.org/10.21236/ADA114514" target="_blank" rel="noopener">https://doi.org/10.21236/ADA114514
Sala, E.和S. Giakoumi, 2018。禁捕海洋保护区是海洋中最有效的保护区。海洋学报75(3):1166-1168。<一个href="https://doi.org/10.1093/icesjms/fsx059" target="_blank" rel="noopener">https://doi.org/10.1093/icesjms/fsx059
Sbragaglia, V., L. Morroni, L. Bramanti, B. Weitzmann, R. Arlinghaus和E. Azzurro。鱼叉捕鱼调节鱼的飞行起始距离:保护、个体大小和携带鱼叉枪的影响。海洋学报75(5):1779-1789。<一个href="https://doi.org/10.1093/icesjms/fsy059" target="_blank" rel="noopener">https://doi.org/10.1093/icesjms/fsy059
斯摩霍恩-韦斯特,P. F., S. E.戈登,A. C.邓普西,S. J. Purkis, S. Malimali, T. Halafihi, P. C. Southgate, T. C. L. Bridge, R. L. Pressey, G. P. Jones. 2019。汤加海洋生态系统管理的社会-环境空间层。太平洋保护生物学27:86-92。<一个href="https://doi.org/10.1071/PC19032" target="_blank" rel="noopener">https://doi.org/10.1071/PC19032
Smallhorn-West, P. F., J. Sheehan, S. Malimali, T. Halafihi, T. C. L. Bridge, R. L. Pressey, G. P. Jones. 2020a。激励共同管理产生影响:推动汤加特别管理区方案在全国成功扩大的机制。保存信件13(6):e12742。<一个href="https://doi.org/10.1111/conl.12742" target="_blank" rel="noopener">https://doi.org/10.1111/conl.12742
史摩霍恩-韦斯特,P. F., K. Stone, D. Ceccarelli, S. MaliMali, T. Halafihi, T. Bridge, R. Pressey, G. Jones。社区管理对沿海渔业资源和生物多样性保护产生积极影响。保存信件13(6):e12755。<一个href="https://doi.org/10.1111/conl.12755" target="_blank" rel="noopener">https://doi.org/10.1111/conl.12755
Tran, D. S. C. K. A. Langel, M. J. Thomas, D. T. Blumstein. 2016。海洋保护区内外未被捕获物种的鱼叉捕鱼行为变化。当前生态62(1):39-44。<一个href="https://doi.org/10.1093/cz/zov006" target="_blank" rel="noopener">https://doi.org/10.1093/cz/zov006
特恩布尔,j.w., e.l.约翰斯顿和G. F.克拉克,2021年。评估部分海洋保护区的社会和生态效益。保护生物学35(3):921 - 932。<一个href="https://doi.org/10.1111/cobi.13677" target="_blank" rel="noopener">https://doi.org/10.1111/cobi.13677
Visconti, P., M. Bakkenes, R. J. Smith, L. jooppa, R. E. Sykes. 2015。全球保护区扩张计划对社会经济和生态的影响。英国皇家学会哲学学报B:生物科学370:20140284。<一个href="https://doi.org/10.1098/rstb.2014.0284" target="_blank" rel="noopener">https://doi.org/10.1098/rstb.2014.0284
Waldron, A., V. Adams, J. Allan, A. Arnell, G. Asner, S. Atkinson, A. Baccini, J. Baillie, A. Balmford,和J. Austin Beau。为自然保护地球30%的土地:成本、收益和经济影响。保育研究所,剑桥大学,英国剑桥。
欢迎R. L.和D. M. Bartley, 1998。目前加强渔业的方法。渔业管理与生态5(5):351-382。<一个href="https://doi.org/10.1046/j.1365-2400.1998.550351.x" target="_blank" rel="noopener">https://doi.org/10.1046/j.1365-2400.1998.550351.x
Zupan, M., e . Fragkopoulou, J. Claudet, K. Erzini, B. Horta e Costa, e . J. Gonçalves。2018.海洋部分保护区:生态效益的驱动因素。生态学报,16(7):381-387。<一个href="https://doi.org/10.1002/fee.1934" target="_blank" rel="noopener">https://doi.org/10.1002/fee.1934

图1


图1所示。本研究包含三个指标和案例的南太平洋地区地图。生产率代表基于单个物种生长速度的新生物量的生产速度(Morais和Bellwood, 2020年)。可捕性表示接近一条暗礁鱼的长矛距离(平均3.37米,范围3.05 - 3.60米)的概率,从而表示捕捞量和努力之间的权衡(Januchowski-Hartley et al. 2014)。脆弱性指的是根据关键生活史特征,鱼类对过度捕捞的易感性(0-100)(张et al. 2007)。每一种管理战略都代表一种可在共同管理安排下采用的方法,尽管这些安排并不相互排斥。部分保护区(绿色)限制了何时、如何或由谁收割。禁捕保护区(红色)是指永久禁止捕鱼的保护区。由于社区和伙伴组织WorldFish之间的保密协议,研究社区在所罗门群岛的确切位置没有被共享。
图2

图2所示。不采储量和PPAs对汤加特别管理区计划目标(a,c)和总(b,d)生产力和生物量的影响。红色和绿色圆圈表示管理区域内的平均值。蓝色圆圈表示开放捕鱼区域的反事实值的平均值,根据9个社会环境变量,横断面在统计上匹配(附录1,表A1, A2)。误差条表示95%的置信区间。星号表示管理区域和预测反事实条件之间的值有显著差异(p < 0.05)。
图3

图3。两种共同管理策略之间刺参科和唇形礁亚科鱼在鱼刺范围内的接近概率差异(附录1,表A5)。捕获概率定义为接近一条暗礁鱼在鱼叉距离(3.37米)内的可能性。灰色圆圈表示在每个鱼科的平均大小下的捕获概率(针叶鱼科22 cm TL, Scarinae 26 cm TL)。按区域类型划分的*表示在p < 0.1处大小对捕获概率有显著影响,在p < 0.05处**表示大小对捕获概率有显著影响。
图4
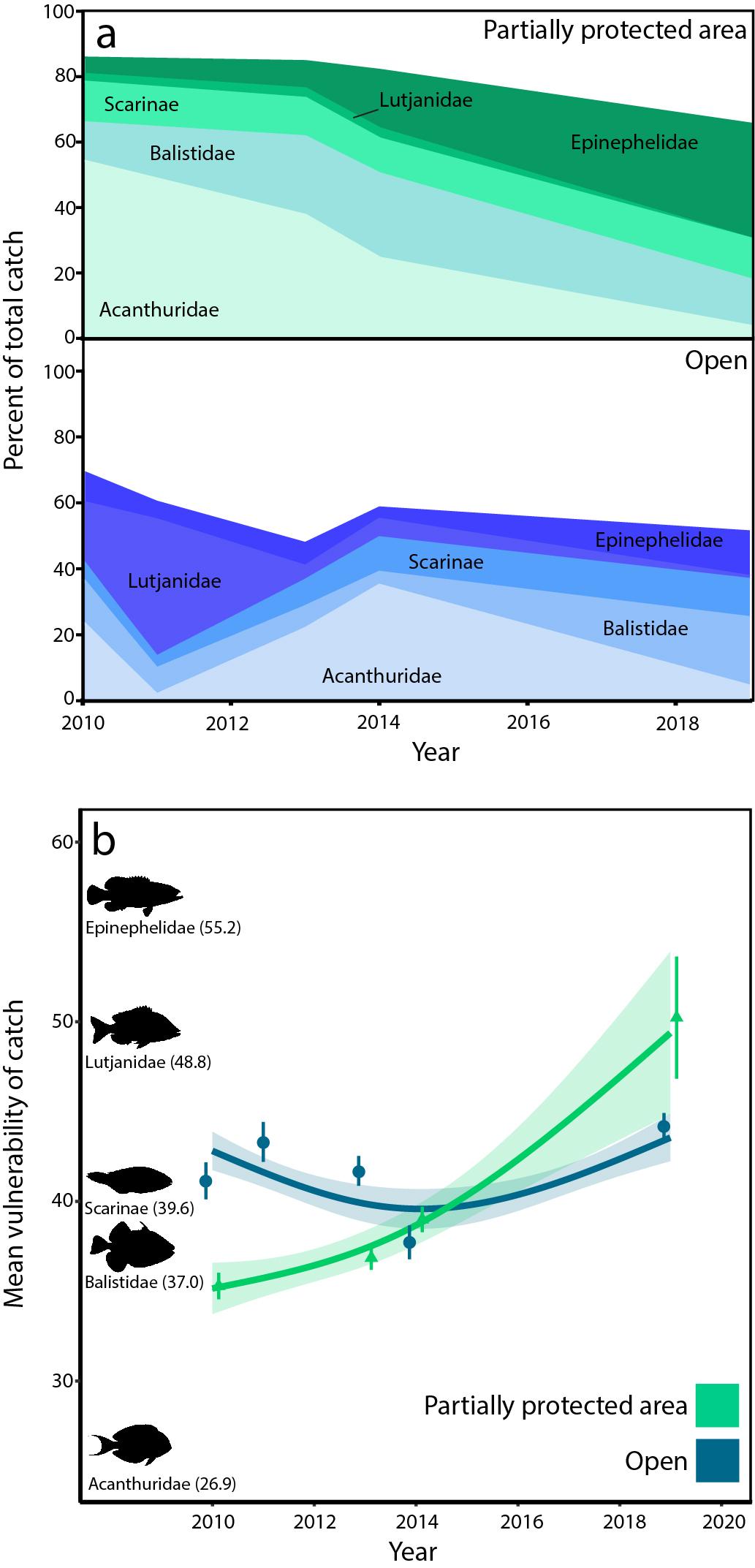
图4。在一个部分保护区(PPAs)的社区捕获成分超过10年。答:在开放和部分受保护的珊瑚礁之间,五种最常见鱼类的渔获量变化百分比。前五大鱼类科的整体渔获率分别为:刺棘科30.5%、刺鲷科15.6%、红鳉科6.6%、灰鲈科8.5%、石斑鱼科9.1%。B:开放和部分受保护的珊瑚礁之间平均生活史脆弱性的变化。绿丝带表示开放期间在周期性收获PPA内捕获的物种的每次捕捞的估计平均脆弱性(±95%置信区间)。蓝丝带代表在社区周围的所有其他珊瑚礁上进行的捕鱼活动的脆弱性。点代表每年(±SE)数据收集的平均值。捕获漏洞的大小从1到100不等,数值越大表示漏洞越大。y轴上的鱼类轮廓表示最常捕捞的前5种鱼类的平均家族水平脆弱性。
表1
表1。南太平洋三个国家关于共同管理对渔业指标的影响的个案研究的详细情况。水下目视测量;部分受保护区域。
| 案例研究 | 国家 | 指示器 | #社区 | 时间 | 数据类型 | 样本大小 | PPA的特点 |
| 1. | 汤加 | 生产力 | 7 | 2016 - 2018 | uv横断面 | 1635年横断面 | 捕鱼的途径和使用只限于社区成员 |
| 2. | 瓦努阿图 | Catchability | 2 | 2011 | 行为调查 | 288条鱼 | 在每6个月预定的3-7天的开放活动期间,只允许社区成员进入和使用 |
| 3. | 所罗门群岛 | 脆弱性 | 1 | 2010 - 2019 | 抓住调查 | 993次 | 在每年2-4周的预先确定的开放活动期间,只有社区成员才能进入和使用钓鱼。 |