






以下是引用这篇文章的既定格式:
lde, D. W., S. M. Cady, R. D. Elmore和S. D. Fuhlendorf, 2022年。木本植物入侵在三个社会和生态多样性的生态区域普遍存在。生态与社会27(3):11。摘要
草原木本植物入侵是全球生物多样性保护面临的最大挑战之一。此外,这是一个社会生态问题,人类的活动和行为导致控制木本植物的生态过程发生了重大变化,而没有充分认识到人类活动的作用,导致世界范围内草地的持续损失。因此,保护专业人员必须了解生态系统、聚落模式和人为开发的碎片化是如何相互作用影响木本植物入侵率的。我们的目标是,首先,描述北美南部大平原三个生态和社会多样性生态区过去20年(2000-2019年)的木本植物入侵率。然后,我们研究了该区域内人为和生物物理变量如何影响入侵率(直接和间接)。结果表明,尽管3个生态区域的社会特征和生态特征存在显著差异,但在研究期间,3个生态区域的木本植物入侵均呈现出持续增加的趋势。这包括堪萨斯州和俄克拉荷马州的弗林特山生态区,该地区经历了广泛和频繁的火灾。在区域尺度上,入侵率与县域平均燃烧面积、树木初始盖度和行行作物破碎度呈直接负相关。开发和行作物农业的覆盖率也通过影响初始树木覆盖率和火灾活动间接改变了一个县的入侵率。木本植物入侵的普遍性质,甚至在经常发生火灾的地区,表明许多草原的管理超出了维持草原和限制木本植物入侵所需的临界生态阈值,这可能对生物多样性产生重大影响。 Our results show that anthropogenic or ecological factors do not act in isolation in their influence on woody plant encroachment and can form complex relationships that shape regional trends in woody plant encroachment.介绍
草原和草原生态系统正经历着从草本物种主导的植物群落向木本植物主导的新状态的快速变化(Roques et al. 2001, Briggs et al. 2005, Stevens et al. 2017)。这种被称为木本植物入侵的制度转变是一项全球性挑战,影响着多个大陆的草原和稀树草原,对全球生物多样性具有重大影响(Stevens等人,2017年,Venter等人,2018年,García Criado等人,2020年)。木本植物的入侵部分归因于人类对火的广泛抑制,这使得对干扰敏感的树种在剩余草原上的覆盖范围和范围增加(Sholes和Archer 1997年,Van Auken 2000年,Bond和Keeley 2005年,Briggs等人2005年,Archer等人2017年)。CO₂含量上升(Bond和Midgley 2000)、氮含量增加(Kochy和Wilson 2001)以及草地过度放牧等因素也与树木侵蚀加速有关(Archer等人,2017年)。草原系统的这些变化会导致生物多样性(Ratajczak et al. 2012)和牧草生产力(Van Auken 2009)的丧失,生态恢复力降低(Alofs and Fowler 2013, Eldridge et al. 2015),生态系统服务中断(Archer et al. 2001, Eldridge et al. 2015, Koch et al. 2015),以及草地依赖物种栖息地的丧失(Grant et al. 2004, Fuhlendorf et al. 2017, Lautenbach et al. 2017)。由于世界各地的草原和稀树草原已经因人类活动而大量减少和退化(Hoekstra et al. 2005),木本植物覆盖的增加对剩余草原的保护构成了生存挑战。了解木本植物入侵的原因和范围是有效应对这一挑战的关键一步。
草原保护的一个新兴范式是,转向理解草原上的木本植物入侵是一个社会生态问题(Donovan等,2018,Luvuno等,2018,Wilcox等,2018)。在这一范式下,人类活动和行为与潜在的生态模式相互作用,以决定景观在草原或林地状态的维持。在大范围内,木本植物入侵受到社会和经济因素的控制,这些因素决定了一个地区是否对木本植物实施有效的控制措施,如频繁的火灾或有针对性的机械/除草剂处理(Wilcox et al. 2018)。这些广泛的模式受到历史土地使用实践(Higgins等人,1999年)、关于土地使用和与使用规定的火相关的责任的地方法规(Yoder等人,2004年,Kreuter等人,2019年)、资本或资源限制以及对火的危险或效用的文化或个人认知(Harr等人,2014年,Hoffman等人,2021年)的影响。与人类活动有关的其他因素,如定居模式、可替代的土地用途和破碎化的强度,可进一步影响树木的侵占,方法是在树木可以生长的地方建立保护区,或改变干扰模式,如火灾和放牧,否则会限制侵占(Hoch和Briggs 1999, Briggs等人2005,Scholtz等人2018a),或通过有意种植树木(Gardner 2009)。然而,降雨(Scholtz et al. 2018b)、土壤(Kaur et al. 2020)和到种子源的距离(Donovan et al. 2018)等生态因素限制了树木的生长位置。社会和生态因素可以进一步相互作用,形成反馈循环,进一步强化侵蚀模式(Ratajczak et al. 2012, Luvuno et al. 2018)。尽管这些已知或感知到的社会和生态景观模式在塑造木本植物分布方面的相互作用,但大多数研究都将这些因素视为独立的过程,很少将它们视为可以形成复杂相互作用和反馈的因果网络的一部分。
北美南部大平原遭受的侵蚀速度是北美其他地区的5到7倍,威胁着该地区大部分剩余的草原(Engle et al. 2008, Barger et al. 2011)。然而,南部大平原是一个文化和生态多样性的地区,这导致许多嵌入的生态区域在上个世纪经历了不同的侵蚀速度(Barger et al. 2011)。东西方向的降水梯度和土壤类型的多样性在很大程度上决定了该地区草原的生产力,进而决定了欧洲定居者的定居模式和土地利用。由此产生的文化和经济做法的差异,使许多生态区域在木本植物入侵和应对方面走上了不同的轨迹(Wilcox等人,2018年)。例如,广泛的灭火、不适当的放牧和行作农业的碎片化促进了对德克萨斯州西南部地区的侵蚀(Box 1967, Diamond and True 2008)。相比之下,弗林特山生态区浅层的岩石土壤阻止了耕作和广泛而持久的火培养,这使得草原得以在低水平的侵蚀下持续存在(Hoy 1989, Briggs et al. 2005),尽管最近的研究表明,木本植物侵蚀可能是该地区一个新出现的问题(Hoch和Briggs 1999, Briggs et al. 2005, Ratajzak et al. 2016)。由于该地区的多样性,南部大平原可能成为了解社会和生态因素如何相互作用影响木本植物入侵率的重要案例。
人们越来越重视将木本植物入侵理解为一种社会生态挑战。为了更好地理解这种全球转变的驱动因素,需要为人为土地利用变化、管理和木本植物入侵率之间的复杂相互作用建立一个清晰的模型。本研究的目的是描述北美南部大平原三个生态区域的木本植物入侵趋势,这三个生态区域的社会和生态组成不同。具体来说,我们测试了这样一个假设:在广泛的社会经济模式支持将广泛和频繁的火灾作为一种管理工具的生态区域,木本植物的入侵会有限,而在对火灾的主流文化态度阻碍了有效的管理实践的生态区域,木本植物的入侵率会最高。我们的次要目标是评估区域人为和生态因素与木本植物入侵之间的因果关系,并确定这些因素如何相互作用形成木本植物入侵模式。为了匹配我们的第一个目标的规模,我们使用与土地使用、发展和碎片化相关的变量作为区域社会和人类活动差异的代理,并使用与火灾和种子源可用性相关的变量来代表生态过程。尽管有许多研究记录了整个大平原木本植物覆盖的增加,以及导致木本植物增加的地方尺度的人为和生态因素,但很少有研究在广泛的区域尺度上检查影响木本植物入侵的因素。通过研究景观水平变量如何影响几个生态区域的木本植物入侵率,这些生态区域构成了生态、人为和生物物理变量的梯度,我们试图为理解大平原的木本植物入侵提供一个框架,这些知识能够指导区域尺度上的保护决策。
研究网站
为了调查木本植物入侵率和影响入侵的因素,我们选择了Wilcox等人(2018)描述的三个生态区域作为案例研究。这些生态区域,德克萨斯州的爱德华兹高原,堪萨斯州和俄克拉何马州的弗林特山,以及俄克拉何马州西部的滚动红山,被选来代表预测影响木本植物入侵率的生态和人为变量的梯度(表1,图1;威尔考克斯等人,2018年)。这三个生态区沿着东西方向的降雨量梯度分布,从东部的弗林特山的年平均降水量约900毫米到红山的670毫米不等。这种降雨梯度导致了从东部的高草草原群落到西部的短草植被群落的生产力梯度(Bragg and Streuter 1996, Assal et al. 2015)。这三个地区都以草地植被群落为主,森林和林地分散,在火灾活动有限的地区持续存在(Wells, 1965, Fowler and Dunlap 1986, Bragg and Streuter 1996)。在欧洲人定居之后,每个地区的土地使用都以放牧为主导,直到20世纪初,定居的增加导致许多地区转向作物生产(Wilcox et al. 2018)。弗林特山是这种作物增产趋势的例外,因为该地区浅层的岩石土壤禁止广泛耕作。在弗林特山区,火的使用主要是作为一种管理工具,用于清除老草落叶和刺激新草生长(Hoy 1989, Briggs et al. 2005)。在滚动红山和爱德华兹高原,使用火作为管理工具的情况要少得多(表1)。尽管这三个生态区的大部分仍然主要用于畜牧活动和农业活动, these regions have also undergone considerable development for human settlements (Table 1). All three ecoregions have similar amounts of total cover for development but show diverging trends in population density and road density (Table 1). The Edwards Plateau contains several metropolitan areas (including San Antonio and Austin, TX) resulting in a higher average population density, although the Rolling Red Hills has remained relatively sparsely populated. The Flint Hills is intermediate in terms of population compared to the other two regions, with several large cities (e.g., Manhattan, KS) interspersed with open rangelands (Hoch and Briggs 1999). Compared to the other two regions, the Flint Hills has a higher overall density of roads per county.
方法
数据准备
侵占率
为了定义所考虑的三个生态区域的范围,我们使用美国环境保护署的III级生态区域来定义爱德华兹高原和弗林特丘陵,使用IV级生态区域来定义Rolling Red Hills。为了划分更小的区域范围,我们将三个生态区域分别划分为县(爱德华兹高原包括36个县,弗林特山包括20个县,滚红山包括9个县)。为了估计木本植物的入侵,我们使用了来自牧场分析平台的数据,年分辨率、30米谷物、植物功能类型数据集(Jones等人,2018;https://rangelands.app/]).对于每个县,我们下载了2000年至2019年树木覆盖的时间序列数据。为了估计每个县树木覆盖的年平均变化率,我们使用了简单线性回归,其中年份(2000-2019年)作为预测变量,每年的估计树木覆盖作为响应变量。估算坡度被解释为每个县每年的平均年变化率,其中正坡度表示木本植物入侵增加,负坡度表示木本植物减少。虽然线性回归不能检测到树木覆盖的非线性关系和短期波动,但使用相对简单的模型使我们能够估计研究期间树木覆盖的总体趋势,同时尽量减少遥感数据固有的方差的影响。
结构方程模型发展与人类生态变量
我们通过回顾评估与木本植物或入侵物种入侵相关的不同因素的近期文献,提出了作为结构方程模型基础的预测(表2)。由于木本植物数据的过程分辨率,我们专注于开发可以由大尺度景观变量表示的假设,如不同土地用途的覆盖、人口密度、以及道路密度,使我们的社会生态数据的分辨率与木本植物数据的分辨率相匹配。
对于人为和生态变量,我们从免费的在线数据源下载了我们所调查的每个地区的县级数据(表1)。因为我们的目标是确定人为与生态模式对木本植物入侵的相对重要性。我们选择关注那些在过去的研究中被证明会影响木本植物入侵的类别变量(表1)。我们计算每个县的人口密度是用2000年人口普查中统计的人口数量除以县的面积(公里²)。我们使用国家TIGER道路数据库计算了道路密度,并计算了每个县每平方公里的道路长度。我们使用来自牧场分析平台的数据来估算研究开始时每个县的树木和草地覆盖。由于“牧场分析”平台不提供对非植被土地覆盖的估计,我们使用2011年全国土地覆盖数据集来估计农业和发展的覆盖率(Scholtz et al. 2018a)。为了找到每个县的年平均降水量,我们使用了气候棱镜组的4公里网格国家天气图,使用了长期平均数据集(https://prism.oregonstate.edu/normals/).对于火灾数据,我们从燃烧严重程度监测趋势(MTBS)数据库(Eidenshink等人,2007年,Picotte等人,2020年)下载了火灾周边形状文件。MTBS数据的最小映射分辨率为404公顷,因此我们的结果可能低估了火灾活动,因为许多小型火灾可能没有被表示出来。然而,由于我们试图在研究期间制定一个火灾活动指数,而不是绘制单个火灾的地图,我们认为MTBS数据是估计大地理区域火灾活动的最佳选择。我们将每个县的所有火灾合并为一个单一的火灾变量,因为我们研究区域记录的75%的火灾被归类为起源不明的火灾。这排除了我们在分析中对规定火与野火的作用提出任何假设。然后,我们估计了该县在2000年至2019年期间每年被烧毁的平均比例。我们将烧毁面积转换为烧毁县的比例,以标准化不同规模县之间的火灾活动度量。为了验证我们对数据集的选择不会影响我们研究的总体结果,我们使用MODIS燃烧面积乘积计算了相同的变量(Roy等人,2005年)。我们发现MTBS数据集估计的平均面积与MODIS数据集的Pearson相关性为r<年代ub>年代= 0.99。这表明数据集有相当多的重叠,而且在我们的研究过程中,两个数据集在很大程度上检测到相同的火灾。所有数据管理和处理都在ArcGIS Pro (ESRI, Redlands, CA)中进行。
数据分析
Ecoregion比较
为了比较这三个生态区域的侵蚀率,我们使用了Kruskal-Wallis检验。然后,我们使用Dunn检验进行事后检验,以确定生态区域之间的两两差异。所有试验均认为p < 0.05水平显著。
Anthropogenic-biophysical变量
利用结构方程建模方法,评价了社会生态因子对木本植物入侵的影响。作为我们分析的基础,我们开发了一个路径图,表示由表2中提出的假设所预测的连接。然后,我们使用R包的最大似然方法评估变量之间关系的强度<年代pan class="ob-q">”lavaan<年代pan class="ob-q">”(Rosseel 2012)。我们没有试图简化原始模型,因为我们正在测试一个关于不同景观变量与木本植物入侵率之间关系的特定假设。我们确实计算了修改指数,以确定是否有任何缺失的因果关系。这就增加了一个县的年平均降水量和树木初始覆盖率之间的单一联系。我们用χ²检验评估模型的最终拟合,并报告自由度和均方根误差(RMSE)。
结果
对土地覆盖数据的分析显示,所有三个生态区域的树木覆盖都有增加的趋势(图2;无花果。爱德华兹高原的树木覆盖率总体变化百分比最低,从2000年到2019年增长了18.62%(表3)。弗林特山生态区的树木覆盖率变化百分比最高,为46.6%,滚动红山生态区居中,为39.2%。由于初始树木覆盖量和生态区域的大小不同,爱德华兹高原、红山和燧石山生态区域分别有372,930 ha、39,896 ha和84,232 ha的草地转化为木本植物(表3)。尽管总体转化为树木的面积存在差异,但三个生态区域在县级上的平均入侵率差异不显著(χ²= 5.00,p = 0.08)。但这可能部分是因为生态区域内木本植物入侵的相当大的变异性(图2;无花果。A1, A2, A3,附录1)。结构方程模型
拟合指数表明我们的模型的整体拟合的混合证据。标准化均方根残差(SRMSR)和比较拟合指数(CFI)拟合良好(SRMSR = 0.048, CFI = 0.96;较低的SRMSR值和接近的CFI值表明拟合良好),而较高的均方根误差(RMSE)值表明大量的方差仍然无法解释(RMSE = 0.187;小于0.06的值被认为是好的;范等人。2016)。我们的模型表明,我们的许多变量都直接或间接地影响着木本植物入侵(图3)。每年被烧毁面积越大的县,木本植物入侵的总体比率越低,支持假设1,即更大的火灾活动将限制树木入侵(β = -0.07, SE = 0.02, p = 0.004)。与假设2预测树木覆盖越大的县的入侵率越高相反,每个县的初始树木覆盖对后续的入侵率有负影响(β = -0.08, SE = 0.03, p = 0.01)。然而,原始树木覆盖率与入侵率之间的关系以及入侵率与全县农作物百分比之间的负相关关系(β = -0.07, SE = 0.03, p = 0.01)为假设3提供了支持,即非草地土地覆盖面积越大的县,入侵率越低。年平均降水量越高,侵蚀率越高,支持假设4 (β = 0.10, SE = 0.04, p = 0.006)。一个县的道路密度和开发覆盖率对木本植物入侵没有显著的直接影响(图3)。
我们还观察到人为变量与木本植物入侵之间的一些间接关系。具体而言,由道路(β = -0.23, SE = 0.15, p = 0.1)或开发(β = -1.11, SE = 0.46, p = 0.02)等替代土地覆盖类型造成的碎片化程度较大的县,其年总燃烧面积较低,而道路密度与燃烧面积呈正相关(β = 1.24, SE = 0.27, p < 0.001)。这为假设5提供了混合支持,假设5指出,碎片化程度越高的县,火灾活动越少。我们发现碎片化程度增加会影响初始树木覆盖的假设(假设6)得到了混合支持,因为农业百分比(β = -0.60, SE = 0.09, p < 0.001)和道路密度(β = -0.79, SE = 0.29, p = 0.10)对初始树木覆盖有负影响,而开发百分比(β = 0.25, SE = 0.32, p = 0.36)对初始树木覆盖没有显著影响。县域人口密度对县域初始树木覆盖(β = 0.34, SE = 0.24, p = 0.15)和平均烧伤面积(β = -0.04, SE = 0.41, p = 0.92)均无影响,表明假说7不受支持。
讨论
从历史上看,降水、土壤和来自食草和火灾的干扰(其中许多是人为的,尤其是在大平原)等因素限制了木本植物在草原上传播和生长的能力(Archer et al. 2017)。由于人类人口已经扩大,并日益成为塑造全球生物群落的主导力量(Ellis和Ramankutty 2008),人类活动的变化在促进全球草原木本植物的增加方面发挥了重要作用(Archer等人,2017年,Stevens等人,2017年,Venter等人,2018年)。在此,我们发现三个以草地为主导的生态区域,它们的社会和生态特征不同,目前正经历着广泛的木本植物侵蚀。最值得注意的是,弗林特山生态区,在那里规定的火是一种常见的草原管理工具,其入侵率与接受有限的火的生态区相当。此外,我们的研究结果表明,人类和生态因素对木本植物入侵的影响很少是孤立的,它们可以形成复杂的关系,影响木本植物入侵的区域趋势。草原是世界上变化最严重的生态系统之一(Hoekstra等人,2005年),木本植物入侵的普遍性质是一项重大的保护挑战,目前的管理实践和战略可能不足以应对(Fogarty等人,2020年,Scholtz等人,2021年)。此外,持续的侵蚀有可能改变剩余草原提供的一些重要的生态系统服务(Archer和Predick 2014年)。未能解释景观变量与木本植物入侵之间的直接和间接关系,会阻碍我们准确预测入侵趋势的能力。
通过提出多个假设,并绘制出描述人为和生态因素直接和间接影响的因果图,我们能够更好地阐明这些变量在形成三个生态区域的木本植物入侵中所起的作用。我们的预测得到了普遍的证实,即在年降水量较大的县木本植物入侵会更高(Sankaran et al. 2005, Scholtz et al. 2018b),而在火灾活动较大的地区木本植物入侵会更低(Archer et al. 2017)。然而,我们发现,其余几个被考虑的变量与木本植物的入侵有更复杂的关系。一些人为变量,如道路和发展,似乎通过其他变量,如烧焦面积,间接影响木本植物的入侵,而其他人为因素,如作物破碎,直接和间接影响木本植物。尽管这支持了之前的研究,即木本植物在碎片化程度较高的地区受到的侵蚀更高(Scholtz 2018a),但这些结果表明,考虑碎片化的具体来源或类型是重要的,因为不同的景观因素可以以各种方式与现有的生态过程相互作用。与我们的假设相反,我们发现在一个县的树木覆盖增加会导致更大的繁殖压力,从而更高的入侵率,在树木覆盖高的县的入侵率较低。缺少对增加的树木覆盖的支持,导致周围草原的入侵率更高,部分原因可能是我们无法区分该地区发现的不同树种:不同的物种在种子产量和入侵能力方面有很大差异(Richardson et al. 1994)。尽管考虑繁殖源和压力可能对评估当地规模的入侵概率很重要(Lockwood等,2005年),但我们的研究结果表明,在区域尺度上,这种关系可能更加复杂。降雨、木本植物种类和新树木入侵的可用空间等因素也可能是重要的考虑因素。
尽管人们越来越认识到木本植物入侵是一个社会-生态过程(Donovan等人,2018,Luvuno等人,2018,Wilcox等人,2018),但这些因素通常被分开研究。人类行为和人为对景观的改造可以通过植树造林或移除树木直接影响木本植物(Gardner 2009, Ding和Eldridge 2019),也可以通过改变生态过程间接影响树木入侵。火灾范围和频率的变化被认为是全球木本植物崛起的一个重要驱动因素(Bond和Keely, 2005年,Archer等人,2017年)。尽管有许多因素可以影响火作为一种管理工具的使用,但我们的研究表明,在研究期间,来自开发和行间作物的碎片化增加与一个县的平均燃烧面积呈负相关,进而可能促进了木本植物的入侵。火灾的减少可能是火灾在整个景观中传播能力发生变化的结果,或因为火灾减少,或在发达地区附近燃烧时使用低强度的火灾(Pickens等人,2017年)。相比之下,在碎片化程度更大的县(尤其是农业),入侵率也更低,这可能是因为树木入侵和生长的草地减少了。这些破碎化和木本植物入侵之间的对比关系突出了综合研究社会和生态因素的重要性,因为单独考虑破碎化的影响可能会导致对景观变化对木本植物入侵的影响的不完全理解。
尽管在管理方法和土地使用方面存在差异,但这三个生态区域的林木入侵率相似。然而,这些相似之处可能是肤浅的,更准确地代表了入侵过程的不同阶段。许多生物入侵遵循一个一致的模式,最初的建立和种群增长可能缓慢,因为繁殖来源有限或与现有物种的相互作用,但一旦达到临界种群规模,就会出现快速的种群增长时期(Crooks 2005),最后随着种群接近景观的承载能力,增长放缓(Shigesada and Kawasaki 1997)。尽管这三个生态区域在历史上都以草地植被为主,但这些生态区域不同的定居和土地利用历史导致了这些区域在我们的研究之前经历了不同程度的树木入侵。弗林特山持续使用规定的火作为一种管理工具,这可能使该地区在入侵的早期阶段保持了下来,在那里,由于频繁的燃烧,新的个体难以建立起来,树木的传播很慢(Hoy 1989, Briggs et al. 2005)。或者,爱德华兹高原可能代表着生物入侵的终点,那里的入侵可能正在放缓,因为该地区已生长树木的数量更多,草地破碎程度更高,限制了新树木生长的可用空间(Archer et al. 2017)。重要的是,尽管目前入侵率相似,但在不同入侵阶段的区域,入侵树木的后续种群动态可能非常不同,管理策略和资源应该在预期这些潜在的种群增长变化的情况下进行配置(Yokomizo et al. 2009, Roberts et al. 2018)。例如,弗林特山的管理应更加强调预防性措施,并在维持该地区草原连通性所需的关键区域有针对性地移除树木,而在爱德华兹高原,可能需要更密集的管理来移除树木,并应将管理目标对准具有最大恢复潜力的区域(Yokomizo等,2009年,Roberts等,2018年)。
尽管森林侵蚀和一个县每年平均燃烧面积之间存在负相关关系,但在广泛流传的火灾文化基本保持的弗林特山生态区,被证明经历着与很少或没有火灾的地区相似的侵蚀率。最近对弗林特山火灾时空模式的研究表明,土地所有者在将火灾应用于景观的频率方面存在很大差异(Ratajzak等人,2016年)。尽管许多土地所有者经常放火,但弗林特山多达57%的草地发生火灾的频率太低,无法防止木本植物的入侵,至少每1至3年发生一次火灾(Ratajzak et al. 2016)。由于其财产的不同土地用途(Coppedge et al. 2001)、个人道德、对不同管理工具的风险感知(Toledo et al. 2013, Harr et al. 2014)、审美偏好(Sharp et al. 2012, Stroman et al. 2020)以及影响其如何管理其财产的资本或资源约束(Toledo et al. 2014),个体土地所有者或机构可能会表现出广泛的行为。此外,许多经理将他们在一年中燃烧的时间改为条件较温和的时期(通常是3月至4月),导致火灾强度较低(Twidwell等人,2016a)。这些较温和的燃烧条件可能会导致缺乏有效控制木本植物所需强度的火灾(Twidwell等人,2016b, Twidwell等人,2019)。另一方面,木本植物入侵的普遍性质,甚至在火灾仍被用作管理工具的地区,也被引用为木本植物入侵的全球驱动因素的证据(Bond和Midgley 2000, Stevens等人2017,García Criado等人2020)。例如,大气CO₂的增加和气候变化导致的温度升高,可能会使利用C₃光合作用途径的木本植物比在许多草原上占主导地位的草本C₄物种更具竞争优势(Bond and Midgley 2beplay竞技000)。了解人类活动对木本植物入侵的原因和后果的关键一步将涉及调查自上而下(如全球气候变化)和自下而上(如火灾或碎片化的局部影响)驱动因素的相对重要性,以及这些驱动因素如何相互作用影响入侵。beplay竞技
我们的研究结果表明,从整体上看,管理者正在输掉与木本植物的战斗。尽管有几十年的刷式管理,数百万美元的成本分摊计划,以及数千名经理的集体努力(Scholtz et al. 2021)。这一问题跨越了地区,这一挑战的规模表明,管理木本植物可能需要重新评估管理的方式和地点。鉴于许多控制木本植物的努力都是短时期的,特别是在这些物种可能已经很常见的地区(Fogarty等人,2021年,Scholtz等人,2021年),可能更好地将资源分配给树木覆盖水平低,但受侵蚀风险较高或具有特别高的保护重要性以防止进一步损失的地区或地方(Yokomizo等人,2009年,Sullins等人,2019年)。为了实现这一目标,管理人员必须了解影响木本植物入侵的景观尺度过程,并制定一个框架,将人为过程和生态过程结合起来,以确定风险地区。我们的工作试图开始解决这一需求。虽然地方过程对于确定木本植物的确切速度和范围无疑是至关重要的,但像我们这样的大范围分析对于指导区域规划是必不可少的。
草地在全球范围内的消失已被描述为迫在眉睫<年代pan class="ob-q">”生物群落的危机<年代pan class="ob-q">”(Hoekstra et al. 2005)和开发承认北美草原多样性和动态特性的模型对于有效分配保护和管理资源至关重要(Wilcox et al. 2018)。面对这一问题,我们需要重新评估我们如何理解影响木本植物入侵的社会和生态因素,以及我们如何将管理实践应用于景观(Twidwell et al. 2013)。目前,弗林特山和红山等生态地区树木覆盖率较低,仍可采取早期干预措施,如增加使用火、除草剂或有针对性地使用机械清除。尽管我们的研究范围很宽,但入侵的区域趋势的变化表明,当地景观因素仍然在决定入侵方面发挥着重要作用(Higgins和Scheiter 2012, Stevens等人2017,Venter等人2018),个体属性可以维持草原和草原生物区系。尽管我们在这项研究中区分了人为变量和生态变量,但在许多情况下,火灾和树木覆盖等变量无法如此明确地定义为与人类行为分离的变量,而聚落模式在决定树木和火灾发生的方式和地点方面发挥着关键作用(Berg et al. 2015)。将木本植物入侵和人类行为结合起来考虑对有效管理至关重要(Wilcox et al. 2018)。然而,与植物群落变化相关的正反馈循环可能很快会加速木本植物的入侵过程,挑战我们用现有工具管理的能力,并可能导致生物多样性不可逆转的损失。
作者的贡献
D. W. L. S. M. C. R. D. E.和S. D. F.设计和构思了这项研究。D. W. L.和S. M. C.进行数据分析。
D. W. L.和S. M. C.撰写手稿。D. W. L. S. M. C. R. D. E.和S. D. F.在稿件准备的各个阶段都提供了反馈和意见。
致谢
这项工作得到了美国农业部国家食品和农业研究所和俄克拉荷马州立大学俄克拉荷马州农业实验站的Hatch OKLO3056项目的支持。我们得到了Bollenbach和Groendyke捐赠野生动物椅的额外支持。
数据可用性
所有使用的数据都可以通过本文描述的在线数据库免费获得。
文献引用
阿洛夫斯,K. M.和N. L.福勒。2013。由于木本植物的入侵,本地草本物种的丧失促进了入侵草的建立。生态94(3):751 - 760。https://doi.org/10.1890/12-0732.1
阿彻,S. R.安德森,K. I. Predick, S. schwin, R. J. Steidl和S. R. Woods. 2017。木本植物入侵:原因与后果。大卫·d·布里斯克主编的25-84页。牧场系统:过程、管理和挑战。施普林格国际,Cham,瑞士。https://doi.org/10.1007/978-3-319-46709-2_2
阿彻,S. R., T. W.布特顿和K. A.希巴德,2001。草原上的树木:木本植物扩张的生物地球化学后果。e - d的115-137页。舒尔茨、S. P.哈里森、M.海曼、E. A.霍兰德、J.劳埃德、I. C.普伦蒂斯和D.希梅尔主编。气候系统中的全球生物地球化学循环。学术,圣地亚哥,加利福尼亚州,美国。
阿彻,S. R.和K. I.预测。2014。灌木丛管理的生态系统服务视角:竞争土地利用目标的研究重点。生态学杂志102(6):1394-1407。https://doi.org/10.1111/1365-2745.12314
阿萨尔,t.j., C. P.梅尔彻和N. B.卡尔,2015。南部大平原生态区域快速评价:预评价报告。报告2331-1258,美国地质调查局,美国弗吉尼亚州莱斯顿。https://doi.org/10.3133/ofr20151003
巴格,N. N., S. R.阿彻,J. L.坎贝尔,C. Y.黄,J. A.莫顿,A. K.纳普。2011。北美旱地木本植物增殖:对生态系统碳平衡影响的综合分析。地球物理研究:生物地球科学116(G4)。https://doi.org/10.1029/2010JG001506
伯格,M. D., M. G. Sorice, B. P. Wilcox, J. P. Angerer, E. C. Rhodes, W. E. Fox, 2015。人口变化驱动了木本植物覆盖趋势——大平原的一个例子。草地生态与管理68(4):315-321。https://doi.org/10.1016/j.rama.2015.05.004
邦德,W. J.和J. E.基利,2005。火作为一种全球“食草动物”:可燃生态系统的生态学和进化。生态与进化动态20(7):387-394。https://doi.org/10.1016/j.tree.2005.04.025
邦德,W. J.和G. F.米格利,2000。草地和稀树草原木本植物入侵的CO₂控制机制。全球变化生物学6(8):865-869。https://doi.org/10.1046/j.1365-2486.2000.00365.x
1967年。德克萨斯州西部的射程恶化。西南历史季刊71:37-45。
布拉格,T. B.和A. A. Steuter. 1996。草原生态学——混合草原。F. B. Samson和F. L. Knopf编辑的53-66页。草原保护:保护北美最濒危的生态系统。美国华盛顿特区岛。
布里格斯,J. M.纳普,J. M.布莱尔,J. L.海斯勒,G. A.霍奇,M. S.莱特,J. K.麦卡伦。2005.转型中的生态系统:中丘草地向灌木林转变的原因和后果。生物科学55(3):243 - 254。https://doi.org/10.1641/0006 - 3568 (2005) 055 (0243: AEITCA) 2.0.CO; 2
科佩奇,B. R., D. M.恩格尔,S. D. Fuhlendorf, R. E.马斯特斯,M. S.格雷戈里,2001。美国南部破碎化大平原草地景观覆盖类型与格局动态景观生态学:677-690。https://doi.org/10.1023/A:1014495526696
《骗子》,J. A. 2005。滞后时间和外来物种:生物入侵的生态学和管理慢动作。Ecoscience 12:316 - 329。https://doi.org/10.2980/i1195-6860-12-3-316.1
Diamond, D. D.和C. D. True, 2008。的分布Juniperus德克萨斯州中部林地与一般非生物站点类型的关系。奥肯主编,48-57页。北美西部Juniperus社区。施普林格,纽约,美国纽约。https://doi.org/10.1007/978-0-387-34003-6_3
丁,J.和D. J.埃尔德里奇。2019。对比去除木本植物对生态系统结构、功能和组成的全球影响。植物生态学、进化与系统学的展望39:125460。https://doi.org/10.1016/j.ppees.2019.125460
多诺万,V. M., J. L.伯内特,C. H.贝尔斯基,H. E. Birgé, R.贝文斯,D. Twidwell和C. R.艾伦。2018。社会-生态景观模式预示着温带草原上原生树种的木质侵蚀。生态学与进化8(19):9624-9632。https://doi.org/10.1002/ece3.4340
德里斯科尔、D. A.、D. Armenteras、A. F. Bennett、L. Brotons、M. F. Clarke、T. S. Doherty、A. Haslem、L. T. Kelly、C. F. Sato和H. Sitters. 2021。火是如何与栖息地丧失和碎片化相互作用的。生物学报,32(3):366 - 366。https://doi.org/10.1111/brv.12687
艾登辛克,施文德,布鲁尔,J。朱,B.奎尔,S.霍华德。2007。一个监测烧伤严重程度趋势的项目。火灾生态学3:3-21。https://doi.org/10.4996/fireecology.0301003
Eldridge, d.j, L. Wang, M. Ruiz‐Colmenero。2015.灌丛侵蚀改变了土壤渗透的空间格局。Ecohydrology 8(1): 83 - 93。https://doi.org/10.1002/eco.1490
Ellis, E. C.和N. Ramankutty. 2008。把人放在地图上:世界的人类生物群落。生态环境科学进展6(8):439-447。https://doi.org/10.1890/070062
恩格尔,D. M., B. R. Coppedge, S. D. Fuhlendorf. 2008。从沙盘到绿色冰川:大平原草原的人类活动与环境变化。o·w·范·奥肯主编,253-271页。北美西部Juniperus《生态学研究》第196卷,施普林格,纽约,美国纽约。https://doi.org/10.1007/978-0-387-34003-6_14
范宇,陈杰,G. Shirkey, R. John, S. R. Wu, H. Park, and C. Shao. 2016。结构方程模型(SEM)在生态学研究中的应用:最新进展。生态过程5:1-12。https://doi.org/10.1186/s13717-016-0063-3
福格蒂,D. T., C.德弗里斯,C.贝尔斯基和D.特威德韦尔,2021。迅速的再次蚕食Juniperus virginiana单次修复治疗后。牧场生态与管理78:112-116。https://doi.org/10.1016/j.rama.2021.06.002
福格蒂,D. T., C. P.罗伯茨,D. R.乌登,V. M.多诺万,C. R.艾伦,D. E.诺尔,M. O.琼斯,B. W.奥尔雷德和D. Twidwell. 2020。木本植物的入侵和优先保育地区的可持续性。可持续性12:8321。https://doi.org/10.3390/su12208321
福勒,N.和D.邓拉普,1986。爱德华高原东部的草地植被。美国米德兰博物学家115(1):146-155。https://doi.org/10.2307/2425844
Fuhlendorf, S. D., T. J. Hovick, R. D. Elmore, A. M. Tanner, D. M. Engle和C. A. Davis. 2017。草原鸡保护中木本植物入侵的层次分析。草地生态与管理70(1):9-14。https://doi.org/10.1016/j.rama.2016.08.010
García克里亚多,M., i.h.迈尔斯-史密斯,A. D.比约克曼,C. E.莱曼和N.史蒂文斯,2020。在气候变化下,苔原和热带稀树草原生物群系的木本植物侵蚀加剧。beplay竞技全球生态与生物地理29(5):925-943。https://doi.org/10.1111/geb.13072
加德纳,R. 2009。建设科技森林:内布拉斯加州沙丘的自然、文化和植树。环境历史14(2):275-297。https://doi.org/10.1093/envhis/14.2.275
格兰特,T. A., E. Madden, G. B. Berkey, 2004。北方混草草原乔灌木入侵对草原鸟类繁殖的启示。野生动物协会通讯32(3):807-818。https://doi.org/10.2193/0091 - 7648 (2004) 032 (0807: TASIIN) 2.0.CO; 2
盖尔巴德,J. L.和J.贝尔纳普。2003。在半干旱地区,道路是外来植物入侵的通道。保护生物学17(2):420-432。https://doi.org/10.1046/j.1523-1739.2003.01408.x
哈尔,R. N., L. W.莫顿,S. R.拉斯克,D. M.恩格尔,J. R.米勒,D.德宾斯基。2014。土地所有者对草地管理风险的认知:木本植物入侵和规定生火。生态与社会19(2):41。https://doi.org/10.5751/ES-06404-190241
希金斯,S. I.和S. Scheiter, 2012。大气CO₂导致植被在局部发生突变,而不是在全球发生突变。自然488:209 - 212。https://doi.org/10.1038/nature11238
希金斯,沙克尔顿,罗宾逊,1999。南非半干旱稀树大草原不同土地利用系统下木质群落结构和组成的变化生物地理学报26(3):619-627。https://doi.org/10.1046/j.1365-2699.1999.t01-1-00317.x
Hoch, G.和J. Briggs. 1999。堪萨斯州北弗林特山东部红雪松的扩张。第十六届北美草原会议论文集(Kearney, 1998)第9-15页。内布拉斯加州大学,科尔尼,内布拉斯加州,美国。
胡克斯特拉,J. M.鲍彻,T. H.里基茨,C.罗伯茨,2005。面对生物群落危机:栖息地丧失和保护的全球差异。生态学通讯8(1):23-29。https://doi.org/10.1111/j.1461-0248.2004.00686.x
霍夫曼,J. K, R. P.比克斯勒,M. L.特德韦尔,L. G.科尔曼,T. W.麦克丹尼尔,U. P.克罗伊特。2021。情感启发式在美国南部大平原私人牧场实施规定火的决策中的影响。社会与自然资源34(5):1-18。https://doi.org/10.1080/08941920.2020.1864534
霍伊,1989。在堪萨斯弗林特山的民间生活中控制牧场燃烧。大平原季刊9(4):231-238。
考尔,R., O.乔希,R. E.威尔,2020年。东部红杉的生态和经济决定因素(Juniperus virginiana)对草原和森林生态系统的侵蚀:一个来自俄克拉荷马州的案例研究。环境管理杂志254:109815。https://doi.org/10.1016/j.jenvman.2019.109815
科赫,B., P. J.爱德华兹,W. U.布兰肯霍恩,T.沃尔特,G.霍弗,2015。灌木侵蚀影响了两个瑞士亚高山牧场上植物、蝴蝶和蚱蜢的多样性。北极、南极和高山研究47(2):345-357。https://doi.org/10.1657/AAAR0013-093
科奇,M.和S. D.威尔逊,2001。北部大平原氮沉积与森林扩张。生态学杂志89(5):807-817。https://doi.org/10.1046/j.0022-0477.2001.00600.x
Kreuter, u P, D. A. Stroman, C. L. Wonkka, J. Weir, A. A. Abney, J. K. Hoffman. 2019。在美国南部平原,土地所有者对使用规定的火的法律责任的认识。牧场生态与管理72:959-967。https://doi.org/10.1016/j.rama.2019.08.004
琼斯,M. O, B. W. Allred, D. E. Naugle, J. D. Maestas, P. Donnelly, L. J. Metz, J. Karl, R. Smith, B. Bestelmeyer和C. Boyd. 2018。牧场监测的创新:1984-2017年美国牧场每年30米植物功能类型百分比覆盖图。生态圈:新兴技术9(9):1-19。https://doi.org/10.1002/ecs2.2430
劳滕巴赫,J. M., R. T.普拉姆,S. G.罗宾逊,C. A.哈根,D. A.豪克斯,J. C.皮特曼。2017。草原景观中小草原鸡对树木的回避。草地生态与管理70(1):78-86。https://doi.org/10.1016/j.rama.2016.07.008
洛克伍德,J. L.凯西和T.布莱克本,2005。繁殖体压力在解释物种入侵中的作用。生态与进化动态20(5):223-228。https://doi.org/10.1016/j.tree.2005.02.004
Luvuno, L., R. Biggs, N. Stevens和K. Esler. 2018。林木入侵是社会生态制度的转变。可持续性10(7):2221。https://doi.org/10.3390/su10072221
莫勒,R. L.和D. G.古丁,2012。绘制2000-2010年堪萨斯州和俄克拉荷马州弗林特山的烧毁区域。大平原研究22(1):15-25。
莫舍,E. S., J. A.西兰德,A. M.拉蒂默,2009。土地利用历史在北美东北部木本植物物种入侵中的作用。生物入侵11(10):2317-2328。https://doi.org/10.1007/s10530-008-9418-8
皮肯斯,B. A., J. F.马库斯,J. P.卡彭特,S.安德森,P. J. Taillie, J. A. Collazo. 2017。城市增长对一种依赖火的鸣禽景观尺度恢复的影响。环境管理学报191:105-115。https://doi.org/10.1016/j.jenvman.2017.01.005
picott, J. J, K. Bhattarai, D. Howard, J. Lecker, J. Epting, B. Quayle, N. Benson和K. Nelson, 2020。对烧伤严重程度程序映射生产程序和数据产品的监测趋势的更改。火灾生态学16:1-12。https://doi.org/10.1186/s42408-020-00076-y
Ratajczak, Z, J. M. Briggs, D. G. Goodin, L. Luo, R. L. Mohler, J. B. Nippert和B. Obermeyer. 2016。评估从高草草原向林地过渡的潜力:我们的操作是否超过了临界火灾阈值?草地生态与管理69(4):280-287。https://doi.org/10.1016/j.rama.2016.03.004
拉塔克扎克,J. B.尼珀特,S. L.柯林斯,2012。木本植物的入侵减少了北美草原和稀树草原的多样性。生态93(4):697 - 703。https://doi.org/10.1890/11-1199.1
理查森,D. M.威廉姆斯,R. J.霍布斯,1994。南半球松树入侵:扩散和入侵性的决定因素。生物地理学报21(5):511-527。https://doi.org/10.2307/2845655
罗伯茨,C. P., D. R.乌登,C. R.艾伦,D. Twidwell. 2018。入侵树的双重思维和规模不匹配极化策略。科学通报13(3):e0189733。https://doi.org/10.1371/journal.pone.0189733
罗克斯,K. G.奥康纳,A. R.沃特金森,2001。非洲稀树大草原灌木入侵的动态:火、草食、降雨和密度依赖性的相对影响。应用生态学报38(2):268-280。https://doi.org/10.1046/j.1365-2664.2001.00567.x
罗塞尔,Y. 2012。lavaan:一个用于结构方程建模的R包。统计软件学报48(2):1-36。https://doi.org/10.18637/jss.v048.i02
Roy, d.p, Y. Jin, P. E. Lewis和c.o . Justice, 2005。利用MODIS时间序列数据进行火灾影响区域系统制图的全局算法原型设计。环境遥感97:137-162。https://doi.org/10.1016/j.rse.2005.04.007
桑卡兰,M., N. P. Hanan, R. J. Scholes, J. Ratnam, D. J. Augustine, B. S. Cade, J. Gignoux, S. I. Higgins, X. Le Roux和F. Ludwig. 2005。非洲稀树大草原林木覆盖的决定因素。自然438:846 - 849。https://doi.org/10.1038/nature04070
斯科尔斯,R.和S.阿彻,1997。热带稀树草原的树-草相互作用。生态系统科学评论28:517-544。https://doi.org/10.1146/annurev.ecolsys.28.1.517
Scholtz, R., S. D. Fuhlendorf和S. Archer. 2018b。气候-火的相互作用限制了北美大平原草原上潜在的木本植物覆盖度和高度。全球生态与生物地理27(8):936-945。
R.肖尔茨,S. D. Fuhlendorf, D. R. Uden, B. W. Allred, M. O. Jones, D. E. Naugle和D. E. Twidwell. 2021。美国南部大平原灌丛治理有效性的挑战。牧场生态与管理77:57-65。https://doi.org/10.1016/j.rama.2021.03.007
R. Scholtz, J. Polo, E. Tanner和S. Fuhlendorf. 2018a。美国南部大平原草地破碎化及其对木本植物覆盖的影响。景观生态学:1785-1797。https://doi.org/10.1007/s10980-018-0702-4
夏普,E. A., P. G.斯普纳,J.米勒和S. V.布里格斯,2012。看不到草,看不到树?澳大利亚东南部的社区价值观和对树木和灌木侵蚀的看法。景观与城市规划33(2):1 - 6。https://doi.org/10.1016/j.landurbplan.2011.11.009
Shigesada, N.和K.川崎,1997。生物入侵:理论与实践。牛津大学出版社,英国牛津。
史蒂文斯,N, C. E.莱曼,B. P.墨菲和G.杜里根。2017。热带稀树草原上的树木被侵占的现象遍及三大洲。全球变化生物学23(1):235-244。https://doi.org/10.1111/gcb.13409
斯特罗曼,D. A.和U. P.克罗伊特。2014。永久保护地役权和土地所有者:评估地役权知识、满意度和伙伴组织关系。环境管理学报146:284-291。https://doi.org/10.1016/j.jenvman.2014.08.007
斯特罗曼,D. A., U. P. Kreuter和C. L. Wonkka。美国南部平原的土地所有者对木本植物的认知和规定了火。科学通报15(9):e0238688。https://doi.org/10.1371/journal.pone.0238688
Sullins, d.s., d.a. Haukos, J. M. Lautenbach, J. D. Lautenbach, S. G. Robinson, M. B. Rice, B. K. Sandercock, J. D. Kraft, R. T. Plumb, J. H. Reitz. 2019。在不同人为影响的景观中保护小草原鸡的策略。生物保护238:108213。https://doi.org/10.1016/j.biocon.2019.108213
托莱多,D. U. P. Kreuter, M. G. Sorice和C. A. Taylor, Jr. 2014。规定燃烧关联在规定火在牧场生态系统应用中的作用。环境管理学报132:323-328。https://doi.org/10.1016/j.jenvman.2013.11.014
托莱多,D. M. G. Sorice和U. P. Kreuter. 2013。社会和生态因素影响人们对应用高强度规定火烧恢复适应火灾的草原生态系统的态度。生态与社会18(4):9。https://doi.org/10.5751/ES-05820-180409
Twidwell, D., W. E. Rogers, S. D. Fuhlendorf, C. L. Wonkka, D. M. Engle, J. R. Weir, U. P. Kreuter, C. A. Taylor. 2013。正在兴起的大平原火灾运动:市民对木本植物入侵的反应。生态学与环境科学进展11(1):e64-e71。
Twidwell, D., W. E. Rogers, C. L. Wonkka, C. A. Taylor Jr, U. P. Kreuter. 2016b。干旱时期的极端规定火降低了木本芽苗的成活率和密度。应用生态学报53(5):1585-1596。https://doi.org/10.1111/1365-2664.12674
Twidwell, D. A. S. West, W. B. Hiatt, A. L. Ramirez, J. T. Winter, D. M. Engle, S. D. Fuhlendorf, J. Carlson. 2016a。植物入侵和防火政策:哪个对高草草原的火灾行为影响更大?生态系统19:356 - 368。https://doi.org/10.1007/s10021-015-9937-y
德威特韦尔,D, c。l。旺卡,h。王,w·e·格兰特,c·r·艾伦,s·d·富伦多夫,a·s·加尔梅斯塔尼,d·g·安吉尔,c·a·泰勒,u·p·克罗伊特。2019。消防管理中的强制性弹性。环境管理学报240:368-373。https://doi.org/10.1016/j.jenvman.2019.02.073
范·奥肯,2000年。北美半干旱草原的灌木入侵。生态学与系统学季刊31:197-215。https://doi.org/10.1146/annurev.ecolsys.31.1.197
范·奥肯,2009。北美西部草原木本植物入侵的原因与后果。环境科学学报30(6):369 - 369。https://doi.org/10.1016/j.jenvman.2009.04.023
文特尔,Z. S., M. D.克莱默,h . j .。霍金斯》2018。木本植物入侵非洲的驱动因素。自然通讯9:2272。https://doi.org/10.1038/s41467-018-04616-8
威尔斯,1965年。大平原地区陡坡林地、输运草地土壤与草地气候概念。科学》148(3667):246 - 249。
威尔科克斯,B. P, A.伯特,S. R.阿彻,S. D. Fuhlendorf, U. P. Kreuter, M. G. Sorice, W. J. van Leeuwen, C. B.邹。2018。从社会生态的角度看木本植物的入侵。生物科学68(9):691 - 705。https://doi.org/10.1093/biosci/biy051
伍兹,N. N., B. L.道斯,E. B.戈尔茨坦,L. J.摩尔,D. R.杨,J. C.津纳特。2019。种子传播与环境过滤的交互作用影响滨海草地林木入侵模式。生态球10 (7):e02818。https://doi.org/10.1002/ecs2.2818
约德,J. D.恩格尔,和S. Fuhlendorf. 2004。生态系统管理的责任、激励和规定火。生态环境科学进展2(7):361-366。https://doi.org/10.1890/1540 - 9295 (2004) 002 (0361: LIAPFF) 2.0.CO; 2
Yokomizo, H. P. Possingham, M. B. Thomas, Y. M. Buckley. 2009。管理入侵物种的影响:了解密度-影响曲线的价值。生态应用19(2):376-386。https://doi.org/10.1890/08-0442.1

图1

图1所示。三个研究生态区域,包括美国北美南部大平原生态区域(灰色)内的弗林特山、滚动红山和爱德华兹高原(用粗体黑线标出的区域)。
图2
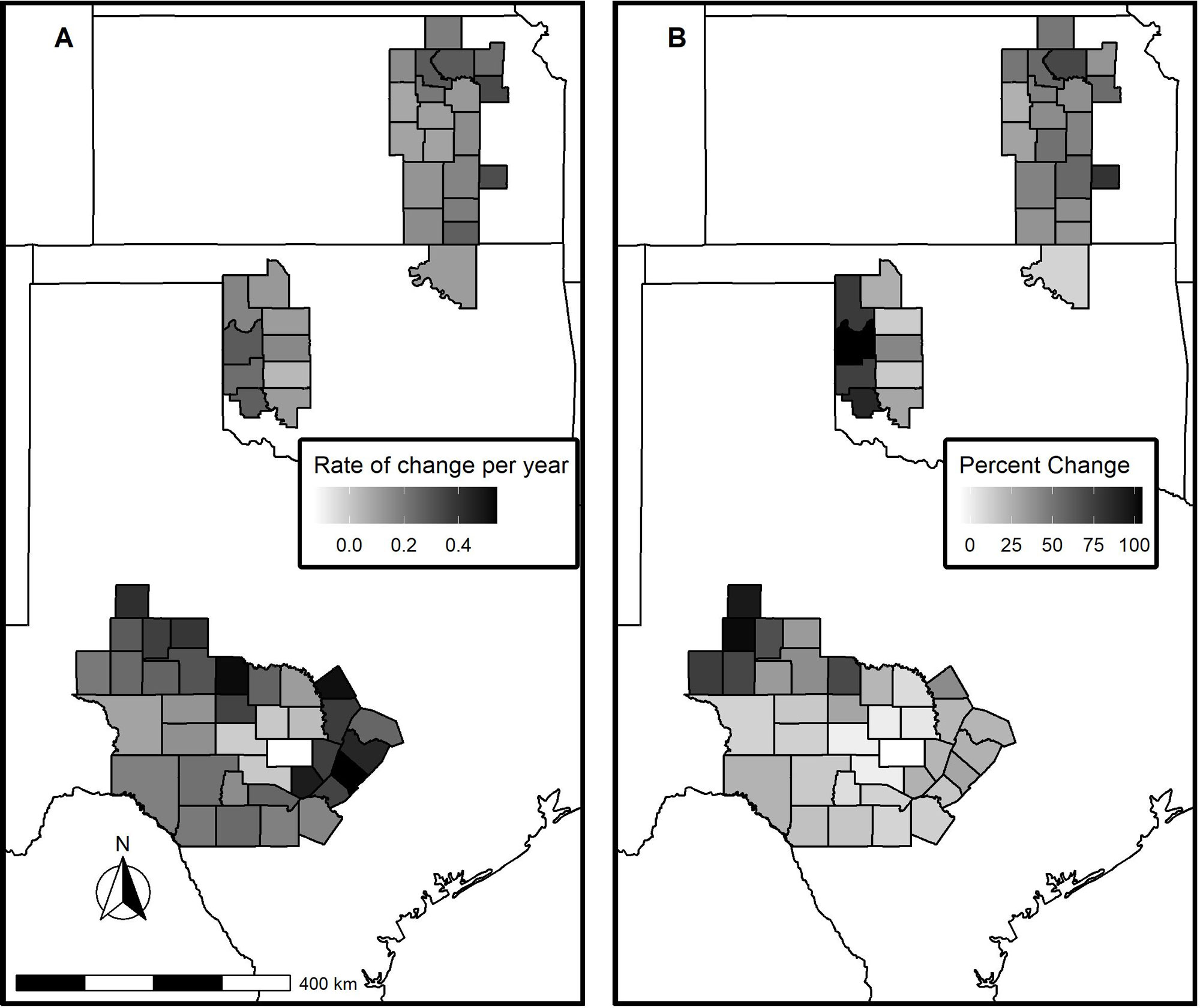
图2所示。美国南部大平原爱德华兹高原、滚动红山和弗林特山生态区的地区,县一级木本植物覆盖度每年的估计变化率(A)和2000年至2019年期间木本植物覆盖度的估计变化率(B)。
图3
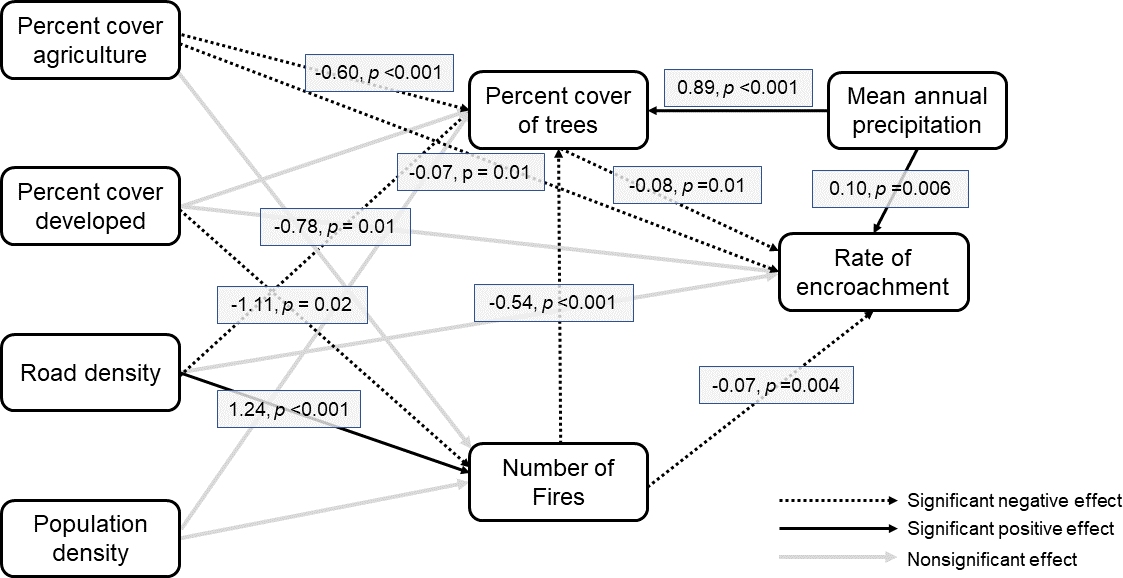
图3。美国南部大平原人为和生物物理变量之间的相互关系和木本植物入侵速率的路径图。显著路径上覆盖的方框表示标准化参数估计和估计的p值。对于非显著路径(灰色线),没有显示估计值和p值。
表1
表1。用于评估2000年至2019年美国南部大平原爱德华兹高原、弗林特山和滚动红山生态区县级木本植物入侵率的人为和生物物理变量。所有变量,平均烧焦面积百分比,代表研究期间(2000-2001年)开始时的条件,因为我们对初始条件将如何影响随后的侵蚀率感兴趣。县级数据的平均值和标准差(括号)显示了每个生态区域。
| 爱德华兹高原 | 燧石山 | 滚滚红山 | 数据源 | |
| 生物物理变量 | ||||
| 平均燃烧面积百分比 | 0.17 (0.35) | 4.73 (4.58) | 0.47 (0.81) | 烧伤严重程度数据库的监测趋势 |
| 覆盖率树木百分比(%) | 24.99 (11.09) | 8.23 (3.68) | 7.13 (3.32) | 牧场分析平台 |
| 年平均降水量(毫米) |
682.59 (142.36) | 905.69 (74.87) | 672.26 (51.85) | PRISM气候数据 |
| 人为的变量 | ||||
| 人口密度(每平方公里人口) | 39 (107.96) | 17.15 (27.73) | 4.71 (3.42) | 2000年的人口普查 |
| 农业覆盖率(%) | 4.9 (7.17) | 32.02 (15.93) | 27.86 (17.15) | 全国土地覆盖数据集 |
| 已开发覆盖率(%) | 5.44 (6.69) | 5.36 (3.07) | 4.63 (0.7) | 全国土地覆盖数据集 |
| 道路密度(每平方公里道路数) | 0.68 (0.85) | 1.39 (0.38) | 0.6 (0.16) | 国家虎路GIS |
表2
表2。描述2000年至2019年美国南部大平原三个生态区木本植物入侵率与人为和生物物理变量之间直接和间接关系的假设。在某些情况下,两个或两个以上的假设为两个变量之间的联系提供了不同的解释,甚至可能有相反的预测(一个假设可能预测积极的影响,而另一个预测消极的影响)。假设用于确定用于后续分析的结构方程模型。
| 直接影响 | 引用 | |
| 假设1 | 经历了更多火灾的县,木本植物的入侵率就会更低,因为火灾会限制新树木的种植。 | 肖尔斯和阿彻1997,布里格斯等人2005,阿彻等人2017 |
| 假设2 | 初始树木覆盖率较高的县有较高的木本植物入侵率,因为这些已经生长的树木可以作为新树木的繁殖源。 | 多诺万等人2018,伍兹等人2019 |
| 假设3 | 由于道路、开发、农业或树木等人为因素造成的草地碎裂,将通过限制树木生长的区域,或由于其他土地用途限制了对树木的管理,从而改变对草地的侵蚀速度。 | Gelbard等人2003,Mosher等人2009,Archer等人2017,Scholtz等人2018b |
| 假设4 | 较高的年降水量将导致较高的侵蚀率,因为林木覆盖潜力已被证明与降水模式有关。 | Sankaran等人2005,Scholtz等人2018a |
间接影响 |
||
| 假设5 | 在草原破碎程度较高的县,火灾活动减少,导致木本植物受到更大的侵蚀。这可能是由于火势蔓延有限,或由于土地使用方式的多样性可能不方便或不允许规定的燃烧。 | Scholtz等人2018b, Donovan等人2020,Driscoll等人2021 |
| 假设6 | 碎片化程度或人口密度较高的县最初的树木覆盖率较高,因为树木被有意种植在人类建筑附近,或者因为不同/不一致的管理做法允许树木在人类建筑周围生长。这可能通过增加繁殖源(更高的入侵)或增加竞争(更低的入侵)来改变入侵率。 | Hoch和Briggs 1999, Gelbard等人2003,Gardner 2009, Scholtz等人2018b |
| 假设7 | 人口密度越高的县,火灾就越少,木本植物受到的侵蚀就越大。这可能是由于使用规定的火、有关烟雾管理的地方法规或可能不方便或不允许规定燃烧的土地使用做法的使用增加了管理者的责任风险。 | Hoch和Briggs 1999, Berg等人2015 |
表3
表3。在研究期开始(2000年)和研究期结束(2019年)期间,美国南部大平原爱德华兹高原、弗林特山和滚动红山生态区的树木覆盖面积(括号中为树木覆盖面积的百分比)。此外,我们还展示了2000年至2019年每个生态区域的树木覆盖率变化百分比。
| 地区 | 初始树木覆盖平方公里 | 最终树木覆盖平方公里 | 变化百分比 |
| 燧石山 | 180770例(6.4%) | 265002例(9.4%) | 46.59% |
| 爱德华兹高原 | 2002122 (26.7%) | 2375052.5 (31.68) | 18.62% |
| 滚滚红山 | 101763.8 (7.7%) | 141659.9 (10.7%) | 39.20% |