






以下是引用本文的既定格式:
周杰,G. P.科菲纳斯,K.基兰,R. B.布恩,L.普吕,K. D. Tape, 2022。beplay竞技气候变化、驼鹿和自给自足的收获:阿拉斯加纽克苏特的社会生态评估。生态与社会27(3):29。摘要
评估气候迅速变暖对北极地区以生存为基础的生计的影响,对于建立农村社区的抵御能力至关重要。我们使用社会-生态系统(SES)框架来评估阿拉斯加北部一个小型土著社区Nuiqsut的驼鹿数量、分布和收获的可能变化范围。我们的结果表明,在村庄用来狩猎的区域内,驼鹿(酒精度酒精度尽管近来为驼鹿提供饲料和掩护的高大灌木有所增加,但驼鹿的数量变化很大。对驼鹿数量的预测也表明未来将继续波动。我们的分析表明,在气候变暖的情况下,未来驼鹿分布的增加将不会出现在猎人可以乘船到达的河流系统中。在气候变暖的情况下,“猎人通道”(即河流通航性)也可能不会增加。因此,努伊克苏特社区提供了一个案例研究,即在变暖条件下,驼鹿栖息地和分布的扩张暴露在较高的环境中,但由于收获方面的限制,对这种变化的敏感性较低。当分别评估社会和生态组成部分时,这些结果并不明显,这说明了SES方法的价值。它们还提供了一个例子,即气候变暖下北极景观和生存资源的迅速变化可能不会转化为收获机会的改变。介绍
北极的快速变暖正在推动社会经济和生态条件的变动性变化(Hinzman等人,2005年,北极理事会2016年,Pürtner等人,2019年)——包括生计收获的变化——这对北极社区在营养和文化上具有重要意义(Kruse 1991年,Burnsilver等人,2016年秋季,Gerlach等人,2017年)。例如,与气候相关的环境条件变化,如水位、冰、雪、侵蚀和风速,都阻碍了北极农村社区的生存旅行和收获能力(Berkes和Jolly 2002, Kofinas等人2010,Hansen等人2013b,布朗等人。2018,Cold等人。2020)。虽然评估气候变化对自给自足的收成的影响对于建立北极社区beplay竞技的抵御能力至关重要(Chapin等人,2004年,McDowell等人,2016年),但我们对许多自给自足物种未来收成的了解有限。
驼鹿(酒精度酒精度)已经扩展到阿拉斯加的北极地区,主要是由于气候变暖引起的高大灌木的扩张(Hall 1973, Tape等人,2016年b).因此,在一些高纬度社区,狩猎驼鹿已成为一种新的机会,这是一种额外的生存资源(Titus等人,2009年,Kofinas等人,2010年)。每头收获的驼鹿可以提供大约244公斤的野生野味(阿拉斯加渔猎部(ADF&G) 2014),在许多社区,驼鹿被认为是一种理想的野生食物。由于预计北极将迅速变暖(Overland等人,2014年,Meredith等人,2019年),驼鹿的栖息地预计将在阿拉斯加北极地区增加两倍以上,驼鹿可能会扩张到靠近许多北极社区的排水地区(Zhou等人,2020年)。然而,尚不清楚这种栖息地和驼鹿分布的增加是否会为阿拉斯加北坡Iñupiat社区提供更大的收获机会。
我们的目标是评估气候变暖对Nuiqsut居民狩猎区驼鹿收获的影响,Nuiqsut是阿拉斯加北坡的Iñupiat社区。我们使用了社会生态系统(SES)框架(Walker和Salt 2006, Chapin等人2009,Collins等人2011)来评估气候变暖对驼鹿生存收获的影响。我们遵循Berman和Kofinas(2004)和Brinkman等人(2013)的观点,将生存资源的可用性定义为资源丰度、资源分布和猎人可及性的组合。基于对Nuiqsut活跃的驼鹿猎人的采访,使用基于代理的模型进行收获模拟(Huston et al. 1988),以及对系统中相互作用的社会和生态组成部分的预期变化的模型预测,我们评估了驼鹿可用性的合理变化。我们进一步评估了在北极变暖导致其他生存资源重大损失的情况下,驼鹿是否可以作为潜在的替代品。由于研究区域的驼鹿栖息地预计将随着变暖而迅速扩大(Zhou et al. 2020),我们假设变暖可能会促使更多驼鹿出现在Nuiqsut附近的河流支流,这将增加收获机会,因此,这可能是气候变暖有利于当地社区的一个案例。
研究区域
我们评估了气候变化对阿拉斯加北坡社区Nuiqsut的猎beplay竞技人未来驼鹿收获的影响(图1)。Nuiqsut是一个Iñupiat村庄,位于科尔维尔河上,距离波弗特海海岸约25公里,主要通过飞机到达。每年的1月至4月都有一条冰路运营,从Nuiqsut到Prudhoe Bay(相距约100公里),在那里它连接到Dalton高速公路。努伊克苏特位于平坦无树的北极海岸平原上,以苔原灌木丛为主,在永久冻土上点缀着许多池塘、浅湖和湿地。研究区内溪流及河流的河岸走廊,生长着以高大灌木为主的植物群落,例如毛毡叶柳树(柳树alaxensis)及西伯利亚桤木(赤杨皮绿冬青ssp。后).在河岸走廊中,毛叶柳是最高和最常见的灌木种类。例如,Zhou et al.(2017)报道称,在调查的河岸走廊中,毡叶柳树占记录灌木的46%,而其他高大灌木物种不到12%。河岸走廊上的毡叶柳斑块是驼鹿的主要栖息地(Mould 1977, Zhou et al. 2017)。高大的灌木和驼鹿主要分布在河流的河岸走廊。小溪和河流通常从南部的布鲁克斯山脉流向北部的波弗特海。科尔维尔河是阿拉斯加北极地区最长(约600公里)、水量最大的河。努伊克苏特采集者使用的驼鹿狩猎区域在8月和9月可以乘船前往,沿着科尔维尔河及其支流前往北极山麓的高大河岸灌木,那里是大多数驼鹿出没的地方(Zhou et al. 2017)。
努伊克苏特居民参与了一种混合的生存-现金经济,其中大多数居民参与了生存经济(ADF&G 2014年,Brubaker等人2014年)。居民们收获了陆地、淡水和海洋物种的混合,包括弓头鲸(Balaena mysticetus)、白鱼(Coregonus种)、北美驯鹿(学家)、驼鹿、海豹(如须、Erignathus barbatus和丝带,Phoca fasciata)、鹅(如白面鹅、雁属albifrons,加拿大,Branta黄花)、北极灰斑鱼(Thymallus arcticus),最少思科(c .沙丁鱼)和波博特(罗大罗大).2014年,纽克苏特居民的人均捕获量估计为406公斤,其中大部分来自弓头鲸、北美驯鹿和鱼类(ADF&G 2014年)。尽管收获量很高,但一份对Nuiqsut家庭的调查报告发现了粮食不安全的程度,38%的家庭没有足够的健康食品,53%的家庭偶尔出现食物不足的情况(Brubaker et al. 2014)。由于航空运输成本高,而且全年都没有公路,村里从商店购买的食物很贵,这凸显了狩猎驼鹿等自给自足的收获对粮食安全的价值。
北坡区(NSB)的人均收入是阿拉斯加最高的,因为它从石油设施基础设施的税收和对居民的转移支付中获得了现金。根据1971年的《阿拉斯加土著索赔解决法案》(ANCSA),阿拉斯加土著地区公司(如北极斜坡地区公司(ASRC))拥有地下权利,而当地村庄公司(如Kuukpik公司)拥有地表权利。ASRC部分拥有Nuiqsut以北13公里的Alpine油田,Kuukpik Corp.与ConocoPhillips签订了地面使用协议。北坡行政区、Kuukpik公司和行政区学区是Nuiqsut的三大雇主。居民也有资格获得阿拉斯加永久基金股息(PFD)、ASRC和Kuukpik Corp.的年度股息。在过去十年中,PFD每年平均每人提供1,390美元(<一个href="https://pfd.alaska.gov/" target="_blank" rel="noopener">https://pfd.alaska.gov/).例如,2018年,持有100股的261名居民每人的股息约为7,000美元(<一个href="http://www.north-slope.org/" target="_blank" rel="noopener">http://www.north-slope.org/).持有Kuukpik Corp. 100股股票的纽克苏特居民每年可获得1.2万至2万美元不等的奖金。然而,失业率很高(例如,美国人口普查局2017年美国社区调查(ACS) 5年调查,2017年为23%),许多居民持有的ASRC或Kuukpik Corp.股票不足100股,有些居民根本没有股票。在1971年以后出生的现有居民(即ANCSA)中,70%以上的人只能通过继承和原始股东的馈赠,或者在极少数情况下,通过现有股东的批准发行新股来拥有股票。随着1971年以后出生的居民越来越多,今后拥有100股的居民有可能会减少。因此,野生食物的可持续收获对于Nuiqsut社区在未来变暖下应对环境快速变化的恢复力至关重要。考虑到它们的体型,驼鹿可以提供大量的野生食物(ADF&G 2014),这可能会使许多现金收入较低的家庭应对这个北极村庄极高的商业食品成本和较低的食品安全(Brubaker et al. 2014)。
方法
概念模型
我们用基于代理的模型(ABM)模拟了灌木丛-驼鹿-猎人系统作为一个社会-生态系统,并评估了变暖下每个关键组成部分未来变化的合理范围。灌木-驼鹿-猎人系统包含了一系列社会和生态成分(图2)。生态成分包括变暖导致的高大灌木栖息地的扩张,这可能会影响驼鹿的分布,在某种程度上,也会影响驼鹿的数量。分析的重点是驼鹿分布和密度的关键变量的变化。社会部分考虑的是进入驼鹿收获区。对可达性的分析主要集中在河流通航性的变化(即进入狩猎区域的可达性)。尽管更好的设备(如喷气船和螺旋桨船)增加了可达性,减少了旅行所需的时间,但河流的通航性被认为主要受到气候引起的水文变化的影响,如河流水位。
我们的概念图(图2)并没有描述影响努伊克苏特居民驼鹿收获的所有变量。例如,对于社会组成部分,控制驼鹿收获的因素的复杂性还包括收获者狩猎的时间、猎人的专业知识、猎人的数量、狩猎法规、狩猎装备、就业和其他产生额外现金的来源(Kruse 1991, Berman和Kofinas 2004)。相反,我们的概念图和模型描述了我们假设气候变化可能影响控制驼鹿收获可用性的关键变量的主要途径。beplay竞技
收集经验数据
为了将驼鹿采伐者的行为和狩猎环境的模型表示参数化,我们与Kuukpik生存监督小组(KSOP)合作,确定并采访了努伊克苏特活跃的驼鹿猎人。2014年,我们对12名活跃的驼鹿收割者进行了半结构化采访。所有参与者都是男性,年龄在31岁至74岁之间,大多数人都有数十年的驼鹿狩猎经验。为了帮助猎人记录地理位置,我们使用了一个显示该地区数字地图的大触摸屏。重要的地理参考点和线,如狩猎场和旅行路线,在具有gis功能的触摸屏上被数字化。访谈数据用于(1)设置模拟环境的地理位置和范围;(2)参数化和校准模拟模型中的狩猎路线、模式、距离、持续时间和选址。我们用数字方式记录了采访的音频和视频。在模拟模型中使用的猎人数量是从ADF&G收集的历史数据中获得的。我们涉及人类受试者的研究已获得阿拉斯加费尔班克斯大学研究所审查委员会(irb# 391916)的批准。在整个研究过程中,我们与KSOP、公众和Kuukpik Corp. (Nuiqsut的村庄公司)举行了多次会议和研讨会,以展示并获得研究计划的批准,给出进展报告,并在最后分享发现并获得额外的反馈。
为了评估驼鹿数量的变化范围,我们使用了1991-2016年ADF&G的驼鹿航空调查数据(n= 26 yr)的游戏管理单元26A,主要位于Colville, Chandler, Anaktuvuk, Killik和Itkillik河的河岸系统。为了估计未来驼鹿种群动态,我们将驼鹿数量作为自回归综合移动平均(ARIMA)模型中的时间序列数据,该模型在数据中寻找模式(Box et al. 2015)。用平均绝对百分比误差(MAPE)评价模型的准确性。为了估计研究区域驼鹿栖息地的未来预测,我们采用了Zhou等人(2020)在政府间气候变化专门委员会(IPCC;beplay竞技Nakicenovic和Swart 2000),在一切照旧的情况下,全球碳排放没有显著减少。为了评估河流通航性的合理变化,我们假设研究区域的水位(河流流量)与降水相关(McClelland et al. 2014)。我们使用了2002-2021年期间月平均排放率的现有数据(n= 20年)来自美国地质调查局(USGS;<一个href="https://www.usgs.gov/" target="_blank" rel="noopener">https://www.usgs.gov/),以及阿拉斯加费尔班克斯大学(<一个href="https://www.snap.uaf.edu" target="_blank" rel="noopener">https://www.snap.uaf.edu).
仿真模型
为了模拟气候变暖下驼鹿收获的变化,我们在NetLogo环境中建立了一个基于空间特异性代理(或个体)的模型(ABM) (Wilensky 1999;NetLogo (<一个href="https://ccl.northwestern.edu/netlogo/" target="_blank" rel="noopener">https://ccl.northwestern.edu/netlogo/))。ABM侧重于智能体与环境的局部交互,以识别突发的一般模式(Huston et al. 1988, Bonabeau 2002)。努伊克苏特驼鹿狩猎区(图1)的模拟环境(2016 × 2010公里)的特点是沿主要河流(Colville、Anaktuvuk、Chandler、Killik和Itkillik河)有适宜的驼鹿栖息地。该模拟的适宜栖息地是基于我们之前的论文(Zhou et al. 2020)估算的驼鹿高灌木栖息地,该论文是基于我们研究地区气候和景观结构变量的Maxent模型开发的。通过Maxent模型(Zhou et al. 2020)计算,特定空间环境中的每个细胞的栖息地适宜性值从0(非栖息地)到1(驼鹿的最佳栖息地)。驼鹿个体在模拟景观上的分布与生境适宜性有关。假设驼鹿最初感知到环境的栖息地质量,然后在合适的栖息地内随机移动。这是一个密度依赖模型,驼鹿避免在一个地点形成大群体。尽管布鲁克斯山脉南部的一些驼鹿是迁徙的(Joly et al. 2015),但我们假设在我们的研究区域没有长距离迁徙。
猎人使用通航水道从纽克苏特前往上游(南部)的驼鹿猎场。在采访中,驼鹿猎人报告说,他们通常可以进入距离河流两岸5公里以内的区域进行捕猎。为了简化模拟模型,我们假设河流两岸的河岸高度(即可达性)没有差异。基于他们的输入,我们假设可达性值在距离河岸5 km处从最高到最低呈线性递减。如果猎人在这个范围内遇到驼鹿,他们就会以成功的概率猎杀驼鹿,这是在模型校准过程中根据观察到的驼鹿收获的验证数据设置的。如果在选定的地点没有收获驼鹿,小组将继续在附近地区搜索,或进一步上游寻找驼鹿,每次狩猎旅行最多10天。在狩猎的第10天,即使猎人没有成功,他们也会回到努伊克苏特。然而,在每年8月1日至9月15日的狩猎季节内,猎人可以进行多次狩猎旅行。模型的一个时间步长对应1 d,在每个模拟年对狩猎持续时间进行模拟。
为了考虑未来驼鹿收获的一系列变化,我们使用了四种情景(表1)<年代ub>0米<年代ub>0)是根据最近的驼鹿种群大小(n= 280)和猎人(n在收割区。通过将收获量的模型输出与可用的历史数据进行比较,用经验数据校准基线情景。我们还运行了只增加驼鹿数量(H<年代ub>0米<年代ub>+),只增加了猎人的数量(H<年代ub>+米<年代ub>0),增加了猎人和驼鹿(H<年代ub>+米<年代ub>+).增加情景的值是研究区域猎人和驼鹿的历史最大观测值(表1)。从基线开始,猎人的数量增加了233%,达到历史最大水平(40个猎人)。驼鹿的密度从平均每1000平方公里9头的基线增加到平均每1000平方公里52头的历史最大值,驼鹿的数量增加了448%。此外,为了评估通航性(通路)对驼鹿收获的影响,我们在河流系统的水流水平和通航性增加的情况下,运行了相同的四种情景。当猎人被允许向上游行进时,驼鹿栖息地内的可到达狩猎区域增加了267%。
为了观察模型的输出,我们记录了每个模拟年收获的驼鹿数量。我们假设一只驼鹿的可食用肉质为244公斤(ADF&G 2014),并将收获的驼鹿数量转换为为Nuiqsut收获的驼鹿质量。将50个模拟年份的平均年总收获与不同的模拟情景进行了比较。
收获的生态系统服务的损失情景
可获得的生存资源的多样化来源有助于北极社区的总体恢复力,在任何特定资源短缺的情况下,为土著社区提供了更换收获物种的选择(Berkes和Jolly 2002, Hansen等人,2013一个).因此,一种资源的收获强度取决于需求,部分取决于其他生存资源的可得性。为了评估驼鹿是否是补偿其他生存资源短缺的潜在替代资源,我们使用了一种简单的方法来评估Nuiqsut对特定资源的依赖程度,并计算了如果该资源因系统遭受灾难性冲击而损失,需要多少驼鹿收获来替代(例如,Kofinas et al. 2016)。我们运行了六个场景,在每个场景中,我们假设一种特定资源(即可食用的肉质量,单位为千克)的总损失,如鲸鱼或驯鹿(表2)<年代pan class="ob-q">”假设的损失<年代pan class="ob-q">”每种资源的百分比(即,<年代pan class="ob-q">”成比例的损失<年代pan class="ob-q">”),并以驼鹿等量(即<年代pan class="ob-q">”麋鹿的单位,<年代pan class="ob-q">”为补充其他生存物种损失的同等数量的肉而需要的驼鹿数量)。在这个情景分析中,我们使用了ADF&G的社区生活信息系统(CSIS;<一个href="http://www.adfg.alaska.gov/sb/CSIS/" target="_blank" rel="noopener">http://www.adfg.alaska.gov/sb/CSIS/).这种情景分析使我们能够考虑到驼鹿在多大程度上可能会填补气候变暖带来的生存短缺。
结果
仿真模型结果
在50年的模拟中,驼鹿数量的增加(H<年代ub>0米<年代ub>+),猎人编号(H<年代ub>+米<年代ub>0),或两者兼有(H<年代ub>+米<年代ub>+)的产量分别使总收获增加了165%、80%和443%(图3)。然而,增加通往排水系统的通航通道导致总收获的收益有限(图3)。例如,当允许猎人进一步上游旅行从而增加狩猎通道时,H .流域的总收获量仅增加17%(即151公斤)<年代ub>0米<年代ub>0场景。
预计驼鹿分布,丰度和收获途径
到2099年,阿拉斯加北极地区的驼鹿栖息地预计将增加一倍以上(Zhou et al. 2020)。然而,将驼鹿分布的空间格局与通航河流系统并置,可以看出气候变暖下驼鹿栖息地空间范围的预期增长可能主要发生在努伊克苏特居民无法通航的河流和溪流沿岸地区(图4)。在可通行的狩猎排水系统内,预计高灌木栖息地的空间范围不会显著增加。其中高大的灌木不太可能从河岸地区扩展到非河岸苔原地区(Tape et al. 2012, Zhou et al. 2020)。
根据狩猎区域内驼鹿种群动态的过去模式,ARIMA模型(准确度= 78%)预测2030年驼鹿数量平均增加到374(±76)头,尽管置信区间很宽(图5)。尽管如此,ARIMA模型预测和未来变暖下驼鹿栖息地的预测增加(Zhou et al. 2020)都表明驼鹿数量在不久的将来可能会增加,但不确定性很高。
对于收获通道,我们发现2002-2020年8月和9月的河流流量(作为水位指标)没有明显的趋势(图6A1, A2)。科尔维尔河的流量模式表明,在2002年至2014年期间,船只进入内陆驼鹿狩猎区并没有增加,2014年这项研究采访的驼鹿猎人证实了这一点。在我们2014年采访后的7年时间里,河流流量持续没有明显的趋势(图6A1, A2)。然而,对于未来的长期趋势,考虑到观测到的流量和降水之间的关系(图6A, B),预测在驼鹿狩猎发生的8月和9月降水量不会显著增加(SNAP n.d.)(图6C)。
生存资源损失的情景
努伊克苏特主要是捕鲸和驯鹿狩猎社区,收获数据显示,该社区高度依赖鲸鱼(例如,占2014年总流入的40%)、驯鹿(占2014年总流入的28%)和鱼类(占2014年总流入的24%)。表2显示了补偿这些主要生存资源的任何假设损失所需的驼鹿收获量。例如,在情景#1中,如果任何灾难性的破坏导致2014年鲸鱼捕获总量减少(67171公斤),Nuiqsut将需要额外的275头驼鹿来维持该社区的维持水平(表2)。如果驯鹿捕获总量减少,Nuiqsut将需要额外的196头驼鹿。目前流入努伊克苏特的驼鹿数量每年不到五头。我们的模拟模型还表明,在历史最高水平1535头驼鹿和40名猎人的情况下,收获系统平均每年只能收获约24(±2)头驼鹿。即使在北极变暖的情况下,驼鹿的栖息地预计会扩大,在不久的将来,驼鹿的收成也不太可能满足潜在的需求。
讨论
北极加速变暖可能会影响生存资源的分布和丰度,以及当地采矿者对资源的获取。利用社会-生态系统(SES)框架(Chapin et al. 2009),我们评估了变暖导致的高大灌木扩张对阿拉斯加高北极地区Nuiqsut未来驼鹿收获的影响。充分描述影响驼鹿收获可用性复杂性的全部变量超出了本文的范围。然而,考虑到我们分析中考虑的变化范围,我们预测,通过高灌木栖息地的变化来调节的北极变暖不太可能增加驼鹿的生存产量。因此,努伊克苏特驼鹿收获的预测变化可以作为一个例子,在这个例子中,快速的景观变化和随之而来的资源分布扩张并没有转化为收获的增加。
我们的研究结果表明,在我们的研究区域(Zhou et al. 2020),驼鹿栖息地的预期快速扩张可能主要发生在北极排水系统中,而这些排水系统大多是猎人乘船无法到达的。在目前可到达的地区,驼鹿的分布没有增加,在预计会有扩大驼鹿栖息地的地区,也没有额外的收获,因此,纽克苏特居民对收获的主要控制似乎是驼鹿的丰富程度,这在未来的趋势中存在很大的不确定性。此外,即使通过逆流而上增加收获机会,收获的额外回报也可能有限。目前,猎人们已经可以进入大部分驼鹿出没的最佳栖息地。布鲁克斯山脉较高的河岸走廊是次优栖息地,高灌木的丰度较低(Zhou et al. 2017)。越来越多的人进入这些贫瘠的栖息地,导致单位劳动收获量的回报递减。
我们认识到气候变化不是直接控制驼鹿数beplay竞技量的唯一因素。自1970年以来,我们研究区域的驼鹿数量一直波动,尽管最近灌木丛扩张,但近年来甚至有所下降(Klimstra和Dagget 2020年)。非气候变量可以强烈影响驼鹿的数量,如捕食(Peterson等人1984年,Gasaway等人1992年,McLaren和Peterson等人1994年,Messier 1994年),疾病(Forbes等人1996年,Nymo等人2016年),寄生虫(Jones等人2019年),营养不良(Kubota等人1970年,O 'Hara等人2001年,Wam等人2018年),密度依赖效应(Seaton等人2011年),以及可能由雪靴兔进行的资源竞争(天兔座也;Dodds 1960, Belovsky 1984, Zhou et al. 2017)。
上述因素也可能与变暖协同影响驼鹿的数量(Weiskopf等人,2019年)。例如,气候变暖也可能影响捕食和资源竞争。局部气候变暖可能影响棕熊(熊属arctos)通过改变当地资源的可用性来提高研究区域的适应性(Hilderbrand等人,2019),这反过来似乎改变了对驼鹿的捕食。变暖引起的灌木扩张也可能通过促进其他灌木依赖物种向北极扩展范围来影响资源竞争(Tape et al. 2016一个, Zhou et al. 2017)。因此,对未来收成影响最大的变量(即驼鹿数量)(图3)也是使用气候预测最难预测的。因此,在这一快速变化的地区,确定驼鹿种群变化的主要驱动因素的未来研究对于理解和管理驼鹿数量和收获机会至关重要。
SES框架的应用强调了分析资源分布和收获获取空间模式对评估气候变化对收获可用性影响的重要性(Brinkman等人,2013年,Brown等人,2015年),这与阿拉斯加土著社区表达的担忧一致(Berman和Kofinas 2004年,北极理事会2016年,Brinkman等人,2016年,Chribeplay竞技stie等人,2018年,Meredith等人,2019年)。如果不考虑狩猎获取的社会成分,评估气候变化对生存资源的影响可能会产生误导和混淆(Gerlach等人,2017年)。beplay竞技来自努伊克苏特的猎人专门使用科尔维尔河及其支流进行秋季驼鹿狩猎,这控制了猎人可以进入的地点和时间。如果没有增强的通航路线,假设装备没有重大变化,猎人就无法进入其他排水系统,在这些系统中,变暖可能会导致驼鹿的栖息地扩张(Zhou et al. 2020)。我们的模拟结果进一步强调了景观猎人可以进入的位置的重要性(Berman和Kofinas 2004, Johnson et al. 2016)。例如,目前猎人们每次猎驼鹿平均要走233公里,据报道每趟要消耗231升汽油(= 305美元)。前往较远的恶劣栖息地成本高昂(Brinkman et al. 2014),收获收益递减。
然而,我们的分析表明,考虑到河流水位的历史模式和流域未来降水的预测,在不久的将来狩猎季节,船只进入不太可能增加。驼鹿狩猎区域的河流流域缺乏大量冰川,因此流域径流不太可能随着变暖而增加(McClelland等人,2014年,Rawlins等人,2019年)。预计的冰冻延迟(Magnuson et al. 2000)可能会延长船只通行时间,但在采访中,猎人表示,8月底和9月初的河流水位通常太低,不适合划船。来自USGS Gauge的数据支持了他们的观察。其他未知因素,如河道和侵蚀的变化,也可能影响某些地点的通航性(Walker和Hudson 2003年,Payne等人,2018年,Stephani等人,2020年),但没有描述北坡大面积地区长期趋势的数据。
采用替代的进入方式,如雪地摩托、全地形车或新的陆路旅行形式,以及法规的改变(如春季狩猎),可能使努伊克苏特驼鹿猎人能够利用相对靠近村庄的排水地区,特别是Colville排水地区西部增加的驼鹿分布(图4C)。然而,在努伊克苏特狩猎区变暖下驼鹿数量的未来轨迹远不确定。北极变暖也使苔原旅行更加危险和不可靠(Brubaker等人2014年,Ford等人2019年,Meredith等人2019年)。
努伊克苏特的驼鹿收获也直接取决于每年参与猎鹿的猎人数量(图3)。驼鹿猎人的历史模式呈稳步下降趋势(附录1:图A1.1A)。如果我们假设村庄人口影响了参与驼鹿收获的猎人总数,那么努伊克苏特人的数量保持稳定,并且基于进出迁移模式,没有猎人增加的迹象(附录1:图A1.1B)。这些模式表明,驼鹿猎人的数量在不久的将来不太可能增加。另一方面,在其他主要生存物种的可得性显著下降的情况下,如果对驼鹿的需求增加,那么猎杀驼鹿的动机和猎人的数量可能会增加。
由于乘船进入河流系统受到限制,我们表明,未来北坡的变暖不太可能让驼鹿弥补快速变化的北极地区其他生存资源的任何可能的重大损失(Hinzman等人,2005年,Kittel等人,2011年,Meredith等人,2019年)。生计系统能够提供给努伊克苏特的驼鹿收获量远远不足以满足潜在的短缺。然而,我们对可能的资源短缺的情景分析没有考虑到社区间的基本粮食共享如何弥补基本粮食收获的潜在短期短缺,这一点在北坡的其他地方得到了证明(Kofinas et al. 2016)。分享食物、装备和劳动力是Iñupiat文化的内在组成部分,也是弹性和适应能力的关键来源(Wenzel 2000, Berkes和Jolly 2002, Ford等人2006,Baggio等人2016,Burnsilver等人2016,Fawcett等人2018)。在Nuiqsut, ADF&G的CSIS报告(ADF&G 2014)估计,2014年,91.4%的社区家庭赠送了收获的食物,96.6%的社区家庭收到了收获的资源。
在过去,不断扩大的驼鹿数量为北部社区提供了重要的生存食物来源(Wolfe和Walker 1987, Titus等人,2009,Kofinas等人,2010)。为了在未来气候变暖的情况下减少自给粮食安全的脆弱性,管理者应重点增加可进入地区的驼鹿收获机会,以提高驼鹿作为Nuiqsut社区自给食物来源的可靠性,特别是在其他自给物种出现短缺和不确定性的情况下(Brinkman et al. 2007, Kofinas et al. 2010, 2016)。
如上所述,驼鹿数量对未来变暖的响应将不是线性的。在变暖条件下,影响驼鹿数量的生物和非生物因素的复杂性不可避免地为预测我们研究地区的驼鹿数量趋势带来了很大的不确定性。为了减少评估驼鹿数量趋势的不确定性,并增加狩猎机会,管理人员可以立即采取一些行动,以增加我们对研究地区预期的未来变暖下驼鹿种群动态的了解。首先,管理者应加大力度监测和评估雪靴兔对潜在资源的竞争及其对驼鹿数量的影响(Tape et al. 2016一个, Zhou et al. 2017)。其次,迫切需要对狼等掠食者进行调查(犬属红斑狼疮)和棕熊,并评估它们对驼鹿数量的捕食影响(Klimstra和Dagget 2020)。最后,对驼鹿栖息地灌木牧草的质量和数量进行调查,以评估未来变暖条件下牧草资源与驼鹿密度的关系。
提高狩猎能力和获取渠道的另一种手段是增加现金收入。在驼鹿狩猎的背景下,需要现金来购买装备(船、帐篷、步枪、冰柜)和天然气,这些都是驼鹿生产赖以生存的必需品。如前所述,石油收入是社区现金投入的主要来源,其未来趋势受到政策决策和市场波动的影响,超出了北坡社区的控制范围。此外,由于这可能会降低现有股东的股票价值和投票权,发行新股的可能性往往较小。不过,Kuukpik Corp.最近投票决定向原股东的直系后代发行50股新股。随着年轻居民份额的增加,Nuiqsut的收获能力可能会增加(Kruse 1991)。
虽然SES框架在用于解决特定环境问题时可能是特殊的和特别的,但一般方法对于评估气候变暖下的收获恢复力是必不可少的(Chapin et al. 2009)。我们证明,使用SES框架评估气候变化对生存收获的影响与单独评估社会和生态系统组成部分的变化得出的结论不同(Chapbeplay竞技in et al. 2006, Liu et al. 2007)。在考虑社会和生态因素如何影响收获机会时,我们的分析表明,在变暖条件下,努伊克苏特社区很可能经历了驼鹿栖息地和分布扩张的高度暴露,但对这种变化的敏感性较低,因为这些生态变化不太可能转化为更大的驼鹿收获。这些发现强调了在评估气候变化对生态系统服务的未来影响时,采用综合和跨学科方法提供全面理解(Berman和Kofinas 2004, Berman等人2004,Kruse等人2004,Chapin等人2009,euuskirchen等人2020)的重要性(Meredith等人2019)。beplay竞技
对本文的回应
欢迎对本文进行回复。如果被接受发表,您的回复将被超链接到文章。<一个href="//www.dpl-cld.com/www/responses.php?articleid=13175&mode=add">要提交回复,请点击此链接.<一个href="//www.dpl-cld.com/www/responses.php?articleid=13175">要阅读已接受的回复,请点击此链接.致谢
我们感谢Nuiqsut和KSOP社区,特别是驼鹿猎人分享他们的关键知识,使这项研究成为可能。我们还要感谢娜奥米·奥尼尔在进行猎人采访时提供的重要帮助。研究期间的财政支持包括Liz Claiborne和Art Ortenberg基金会,UAF论文完成奖学金,UAF Jessie O'Bryan McIntosh奖学金,阿拉斯加EPSCoR NSF奖#OIA-1757348,以及阿拉斯加州,国家社会环境综合中心(SESYNC), ARCUS和国家气候适应科学中心。阿拉斯加EPSCoR NSF奖# OIA-1208927和NSF奖# 1263854授予G.P.K, NSF奖#1833056授予K.D.T.
数据可用性
支持本研究结果的数据/代码可以在“figshare”中公开获取<一个href="https://doi.org/10.6084/m9.figshare.14443694" target="_blank" rel="noopener">https://doi.org/10.6084/m9.figshare.14443694https://doi.org/10.6084/m9.figshare.14443373https://doi.org/10.6084/m9.figshare.14443376https://doi.org/10.6084/m9.figshare.14443505.阿拉斯加大学费尔班克斯研究所审查委员会(#391916)批准了这项研究的伦理批准。
文献引用
阿拉斯加渔猎局(ADF&G)。2014.阿拉斯加渔猎部的社区生存信息系统。阿拉斯加渔猎部,安克雷奇,阿拉斯加,美国。URL:<一个href="http://www.adfg.alaska.gov/sb/CSIS/" target="_blank" rel="noopener">http://www.adfg.alaska.gov/sb/CSIS/
北极理事会,2016。北极恢复力报告。M. Carson和G. Peterson,编辑,第1-218页。斯德哥尔摩环境研究所和斯德哥尔摩恢复力中心,斯德哥尔摩,瑞典。
巴乔,J. A., S. B. BurnSilver, A. Arenas, J. S. Magdanz, G. P. Kofinas, M. De Domenico, 2016。多元社会生态网络分析揭示了社会变迁对社区稳健性的影响大于资源消耗。中国科学院学报(自然科学版)43(6):553 - 553。<一个href="https://doi.org/10.1073/pnas.1604401113" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.1604401113
别洛夫斯基,1984。驼鹿与雪靴兔的竞争与觅食理论的机械解释。环境科学,61(2):150 - 159。<一个href="https://doi.org/10.1007/BF00396753" target="_blank" rel="noopener">https://doi.org/10.1007/BF00396753
贝尔克斯,F.和D.乔利,2002。适应气候变化:加拿大西部beplay竞技北极社区的社会生态恢复力。生态与社会5(2):18。<一个href="https://doi.org/10.5751/ES-00342-050218" target="_blank" rel="noopener">https://doi.org/10.5751/ES-00342-050218
伯曼,M.和G.科菲纳斯,2004。狩猎模式:基于理性的选择方法,分析气候对北极社区生存狩猎的影响。生态经济49(1):31-46。<一个href="https://doi.org/10.1016/j.ecolecon.2003.12.005" target="_blank" rel="noopener">https://doi.org/10.1016/j.ecolecon.2003.12.005
伯曼,M., C.尼科尔森,G.科菲纳斯,J. O. E.特特里奇和S.马丁,2004。北极小社区的适应性和可持续性:基于代理的模拟模型的结果。北极57(4):401 - 414。<一个href="https://doi.org/10.14430/arctic517" target="_blank" rel="noopener">https://doi.org/10.14430/arctic517
波纳波,2002年。基于代理的建模:模拟人类系统的方法和技术。中国科学院学报99(3):7280-7287。<一个href="https://doi.org/10.1073/pnas.082080899" target="_blank" rel="noopener">https://doi.org/10.1073/pnas.082080899
鲍克斯,g.e.p, g.m.詹金斯,g.c.r insel, g.m.l jung, 2015。时间序列分析:预测与控制。第五版。威利,霍博肯,新泽西州,美国。
布劳德,S. R. 2010。努伊克苏特,卡克托维克和巴罗的生存地图。为美国内政部阿拉斯加OCS地区矿产管理处准备的报告。Stephen R Braund and Associates,安克雷奇,阿拉斯加,美国。
布林克曼,t.j., W. D.汉森,F. S.查平,G.科菲纳斯,S.伯恩西尔弗,T. S.鲁普,2016。北极社区认为气候对获取的影响是对生存资源可用性的重大挑战。气候变化39(4):457 - 457。<一个href="https://doi.org/10.1007/s10584-016-1819-6" target="_blank" rel="noopener">https://doi.org/10.1007/s10584-016-1819-6
布林克曼,T. J., G.科菲纳斯,W. D.汉森,F. S.查平,III .和S.鲁普,2013。一个管理狩猎的新框架:为什么我们应该把重点从丰富转向可得性。野生动物专业人士7(3):38-43。
T.布林克曼,J.凯利,M.凡戴克,A.菲尔曼,A.斯普林斯汀,2014。燃料成本对高纬度生存活动的影响。生态与社会19(4):18。<一个href="https://doi.org/10.5751/ES-06861-190418" target="_blank" rel="noopener">https://doi.org/10.5751/ES-06861-190418
T.布林克曼,G.科菲纳斯,F. S.查平和D.珀森,2007。猎人适应性对自给狩猎系统恢复力的影响。生态人类学杂志11(1):58-63。<一个href="https://doi.org/10.5038/2162-4593.11.1.4" target="_blank" rel="noopener">https://doi.org/10.5038/2162-4593.11.1.4
布朗,C. L., K. A.西顿,T. J.布林克曼,E. S.尤斯基琛,K.基兰。2015。弹性理论在阿拉斯加驼鹿猎人系统管理中的应用。生态与社会20(1):16。<一个href="https://doi.org/10.5751/ES-07202-200116" target="_blank" rel="noopener">https://doi.org/10.5751/ES-07202-200116
布朗,D. R. N, T. J.布林克曼,D. L.韦尔比拉,C. L.布朗,H. S.科尔德,T. N.霍林沃斯,2018。改变河冰的季节性和对阿拉斯加内陆社区的影响。天气,气候,社会10(4):625-640。<一个href="https://doi.org/10.1175/WCAS-D-17-0101.1" target="_blank" rel="noopener">https://doi.org/10.1175/WCAS-D-17-0101.1
布鲁贝克,M., J.贝尔,H.丁曼,M.伊塔,K.卡萨克,2014。beplay竞技阿拉斯加纽克苏特的气候变化,社区健康战略。阿拉斯加土著部落健康联盟(ANTHC),安克雷奇,阿拉斯加,美国。URL:<一个href="https://anthc.org/wp-content/uploads/2016/01/CCH_AR_072014_Climate-Change-in-Nuiqsut.pdf" target="_blank" rel="noopener">https://anthc.org/wp-content/uploads/2016/01/CCH_AR_072014_Climate-Change-in-Nuiqsut.pdf
伯恩西尔弗,S., J.马格丹兹,R.斯托茨,M.伯曼,G.科菲纳斯,2016。混合经济是持久的还是过渡性的?来自阿拉斯加北极地区的社交网络证据。美国人类学家118(1):121-129。<一个href="https://doi.org/10.1111/aman.12447" target="_blank" rel="noopener">https://doi.org/10.1111/aman.12447
查平,F. S., M.霍尔,S. R.卡朋特,J.卢布琴科,B.沃克,T. V .卡拉汉,C.福尔克,S. A.莱文,k.g.。Mäler, C. Nilsson, S. Barrett, F. Berkes, A.-S。Crépin, K. Danell, T. Rosswall, D. Starrett, A. Xepapadeas和S. A. Zimov. 2006。建立抵御能力和适应能力,以应对北极变化。中35(4):198 - 202。<一个href="https://doi.org/10.1579/0044-7447(2006)35%5B198:BRAATM%5D2.0.CO;2" target="_blank" rel="noopener">https://doi.org/10.1579/0044 35 - 7447 (2006) [198: BRAATM] 2.0.CO; 2
查宾,F. S., G. P.科菲纳斯,C.福克,编辑。2009.生态系统管理原则:变化世界中基于弹性的自然资源管理。Springer-Verlag,纽约,美国。<一个href="https://doi.org/10.1007/978-0-387-73033-2" target="_blank" rel="noopener">https://doi.org/10.1007/978-0-387-73033-2
查平,F. S., G.彼得森,F.贝尔克斯,T. V.卡拉汉,P.安格尔斯塔姆,M.普斯,C.贝尔,Y.伯杰隆,a . S。克雷平,K.达内尔,T.埃尔mqvist, C.福尔克,B.福布斯,N.弗雷斯科,G.朱代,J.尼梅拉,A.什维年科,G.怀特曼,2004。北方地区对社会和环境变化的恢复力和脆弱性。中33(6):344 - 349。<一个href="https://doi.org/10.1579/0044-7447-33.6.344" target="_blank" rel="noopener">https://doi.org/10.1579/0044-7447-33.6.344
克里斯蒂,K. S., T. E.霍尔曼,H. P.亨廷顿,J. R.洛夫沃恩,2018。以传统生态知识为依据的结构化决策分析是在不断变化的北极加强生存系统的工具。生态与社会23(4):42。<一个href="https://doi.org/10.5751/ES-10596-230442" target="_blank" rel="noopener">https://doi.org/10.5751/ES-10596-230442
霍林沃斯、T. J.布林克曼、C. L.布朗、T. N.霍林沃斯、D. R. N.布朗、K. M.海林加。评估阿拉斯加内陆地区生存旅行对环境变化影响的脆弱性。生态与社会25(1):20。<一个href="https://doi.org/10.5751/ES-11426-250120" target="_blank" rel="noopener">https://doi.org/10.5751/ES-11426-250120
柯林斯,S. L., S. R.卡朋特,S. M.斯文顿,D. E.奥伦斯坦,D. L.奇尔德斯,T. L.格拉森,N. B.格林,J. M.格罗夫,S. L.哈兰,J. P.凯伊,A. K.纳普,G. P.科菲纳斯,J. J.马格努森,W. H.麦克道尔,J. M.梅拉克,L. A.奥格登,G. P.罗伯逊,M. D.史密斯,A. C.惠特默。2011。长期社会生态研究的综合概念框架。生态环境科学进展9(6):351-357。<一个href="https://doi.org/10.1890/100068" target="_blank" rel="noopener">https://doi.org/10.1890/100068
多兹,d.g, 1960。纽芬兰驼鹿和雪靴兔的食物竞争和范围关系。哺乳动物杂志24(1):52-60。<一个href="https://doi.org/10.2307/3797356" target="_blank" rel="noopener">https://doi.org/10.2307/3797356
尤斯基兴、e.s.、K.提姆、A. L.布林、S.格雷、T. S.鲁普、P.马丁、J. H.雷诺兹、A.塞塞尔、K.墨菲、J. S.利特尔、A.班尼特、W. R.博尔顿、T.卡曼、H.热内、B.格里菲斯、T.库尔考斯基、M. J.劳拉、S.马尔琴科、D.尼克斯基、S.潘达、V.罗曼诺夫斯基、R.鲁特、C. L.塔克和A. D.麦奎尔。2020.共同生产知识:阿拉斯加北极地区资源管理的综合生态系统模型。生态环境科学进展18(8):447-455。<一个href="https://doi.org/10.1002/fee.2176" target="_blank" rel="noopener">https://doi.org/10.1002/fee.2176
秋季,J. A. 2016。当代阿拉斯加鱼类和野生动物收成的区域格局。北极69(1):47 - 64。<一个href="https://doi.org/10.14430/arctic4547" target="_blank" rel="noopener">https://doi.org/10.14430/arctic4547
弗西特,D., T.皮尔斯,R.诺泰纳,J. D.福特,P.科林斯,2018。西北地区乌鲁克哈托克的因纽特人对不断变化的环境条件的适应能力。极地记录54(2):119-132。<一个href="https://doi.org/10.1017/S003224741800027X" target="_blank" rel="noopener">https://doi.org/10.1017/S003224741800027X
L. B.福布斯,S. V.泰萨罗和W.李斯,1996。实验研究流产布鲁氏菌在慕斯(酒精度酒精度).野生动物学报32(1):94-104。<一个href="https://doi.org/10.7589/0090-3558-32.1.94" target="_blank" rel="noopener">https://doi.org/10.7589/0090-3558-32.1.94
福特,J. D.克拉克,T.皮尔斯,L.贝朗-福特,L.科普兰,J.道森,M.纽和S. L.哈珀,2019。改变了北极社区对冰、土地和水的获取。自然气候变化9beplay竞技(4):335-339。<一个href="https://doi.org/10.1038/s41558-019-0435-7" target="_blank" rel="noopener">https://doi.org/10.1038/s41558-019-0435-7
福特,J. D.斯密特,J.万德尔,2006。北极对气候变化的脆弱性:来自加拿大beplay竞技北极湾的案例研究。全球环境变化16(2):145-160。<一个href="https://doi.org/10.1016/j.gloenvcha.2005.11.007" target="_blank" rel="noopener">https://doi.org/10.1016/j.gloenvcha.2005.11.007
W. C.加萨威,R. D.波尔杰,D. V.格兰加德,G.大卫,R. O.斯蒂芬森和D. G.拉森。1992。捕食在阿拉斯加和育空地区限制低密度驼鹿的作用以及对保护的影响。野生动物专论120:3-59。
格拉赫,S. C., P. A.洛林,G.科菲纳斯,H.佩恩,2017。白令海、波弗特和楚科奇海社区对快速变化的适应能力。L. D.欣茨曼、P.奥特里奇、A.克莱皮科夫、J. E.沃尔什、M.阿纳尼切娃、T.阿姆斯特朗、J.本特森、G.弗莱托、S. C.格拉赫、H. P.亨廷顿、G.科菲纳斯、P.洛林、A. L.洛夫克拉夫特、L. eerks - medrano、E.尼基蒂娜、B. L.普雷斯顿、S. F.特雷纳、J.甘伯和L.索尔斯坦森,编辑。变化中的北极适应行动:来自白令海/楚科奇/波福特地区的观点。北极监测和评估计划技术报告,挪威奥斯陆。<一个href="https://doi.org/10.2172/1435018" target="_blank" rel="noopener">https://doi.org/10.2172/1435018
霍尔,1973年。阿拉斯加北部驼鹿活动范围扩大的考古和最新证据。哺乳动物杂志54(1):294-295。<一个href="https://doi.org/10.2307/1378902" target="_blank" rel="noopener">https://doi.org/10.2307/1378902
韩森,T. J.布林克曼,F. S.查平,C.布朗。满足土著的生存需求:阿拉斯加农村猎物转换的案例。野生动物的人类维度18(2):109-123。<一个href="https://doi.org/10.1080/10871209.2012.719172" target="_blank" rel="noopener">https://doi.org/10.1080/10871209.2012.719172
汉森,W. D., T. J.布林克曼,M. Leonawicz, F. S. Chapin, G. P. Kofinas. 2013b。阿拉斯加北坡每日风速的变化:对农村狩猎机会的影响。北极66(4):448 - 458。<一个href="https://doi.org/10.14430/arctic4331" target="_blank" rel="noopener">https://doi.org/10.14430/arctic4331
希尔德布兰德,g.v, K. Joly, M. S. Sorum, M. D. Cameron和D. D. Gustine。2019。棕熊(熊属arctos) 1977-2016年北极地区的体型、状况和生产力。极地生物学42(6):1125-1130。<一个href="https://doi.org/10.1007/s00300-019-02501-8" target="_blank" rel="noopener">https://doi.org/10.1007/s00300-019-02501-8
Hinzman, l D。:D . Bettez w·r·博尔顿f·s·查宾,m . b . Dyurgerov c . l . Fastie b·格里菲斯r·D·霍利斯特a .希望惠普亨廷顿,a . m .詹森·g·j·贾,t·乔根森D·l·凯恩·D·r·克莱因g . Kofinas a·h·林奇a·h·劳埃德·a·D·麦奎尔f·e·纳尔逊w . c . Oechel t . e . Osterkamp c·h·拉辛v . e . Romanovsky r . s .石头,D . a . Stow m . Sturm c . e . Tweedie g . l . Vourlitis m·D·沃克·D·a·沃克·j·韦伯j·m·威尔克k . s .睫毛吉川,2005。阿拉斯加北部和其他北极地区最近气候变化的证据和影响。beplay竞技气候变化72(3):251-298。<一个href="https://doi.org/10.1007/s10584-005-5352-2" target="_blank" rel="noopener">https://doi.org/10.1007/s10584-005-5352-2
M.休斯顿,D.迪安吉利斯和W.波斯特,1988。新的计算机模型统一了生态理论。生物科学38(10):682 - 691。<一个href="https://doi.org/10.2307/1310870" target="_blank" rel="noopener">https://doi.org/10.2307/1310870
约翰逊,I., T.布林克曼,K.布里顿,J.凯利,K.亨特马克,B.莱克和D.韦尔比拉。2016。量化阿拉斯加农村猎人的获取。野生动物的人类维度21(3):240-253。<一个href="https://doi.org/10.1080/10871209.2016.1137109" target="_blank" rel="noopener">https://doi.org/10.1080/10871209.2016.1137109
乔利,K. T.克雷格,M. S.索伦,J. S.麦克米兰,M. A.斯平德勒,2015。阿拉斯加中北部上科尤库克河流域的驼鹿运动模式。酒精度51:87 - 96。
琼斯,H., P. Pekins, L. Kantar, I. Sidor, D. Ellingwood, A. Lichtenwalner和M. O 'neal。2019.驼鹿死亡率评估(酒精度酒精度)在连续几年的冬天里的幼崽(矩头蝉属albipictus)新罕布什尔州和缅因州(美国)的动物流行病。动物科学进展(1):39 - 44。<一个href="https://doi.org/10.1139/cjz-2018-0140" target="_blank" rel="noopener">https://doi.org/10.1139/cjz-2018-0140
T. G. F.基特尔,B. B.贝克,J. V.希金斯,J. C.哈尼,2011。阿拉斯加北坡生态系统和景观的气候脆弱性。区域环境变化11;1): 249 - 264。<一个href="https://doi.org/10.1007/s10113-010-0180-y" target="_blank" rel="noopener">https://doi.org/10.1007/s10113-010-0180-y
克里姆斯特拉,R.和C. T.达格特,2020年。驼鹿管理报告和计划,游戏管理单元26A:报告期间为2010年7月1日至2015年6月30日,计划期间为2015年7月1日至2020年6月30日。物种管理报告和计划ADF&G/DWC/SMR&P-2020-9,阿拉斯加渔猎部,美国阿拉斯加州朱诺。
Kofinas, G., S. B. Burnsilver, J. Magdanz, R. Stotts, M. Okada, 2016。生活共享网络和合作:卡克托维克,温赖特,和威尼蒂,阿拉斯加。BOEM报告2015-023DOI;AFES报告MP 2015-02。阿拉斯加费尔班克斯大学自然资源与推广学院,阿拉斯加,美国。
科菲纳斯,G. P., F. S.查平,S. BurnSilver, J. I.施密特,N. L. Fresco, K. Kielland, S. Martin, A. Springsteen, T. S. Rupp, 2010。阿萨巴斯坎人生存系统对阿拉斯加内陆气候变化的恢复力。加拿大森林研究40(7):1347-1359。<一个href="https://doi.org/10.1139/X10-108" target="_blank" rel="noopener">https://doi.org/10.1139/X10-108
克鲁斯,1991。阿拉斯加因纽皮亚人的生存和工资就业模式:理解个人选择。人类组织50(4):317-326。<一个href="https://doi.org/10.17730/humo.50.4.c288gt2641286g71" target="_blank" rel="noopener">https://doi.org/10.17730/humo.50.4.c288gt2641286g71
Kruse, J. A., R. G. White, H. E. Epstein, B. Archie, M. Berman, S. R. Braund, F. S. Chapin, J. Charlie, C. J. Daniel, J. Eamer, N. Flanders, B. Griffith, S. Haley, L. Huskey, B. Joseph, D. R. Klein, G. P. Kofinas, S. M. Martin, S. M. Murphy, W. Nebesky, C. Nicolson, D. E. Russell, J. Tetlichi, A. Tussing, m.d. Walker, O. R. Young. 2004。北极社区的可持续性建模:研究人员和当地知识持有者的跨学科合作。生态系统7(8):815 - 828。<一个href="https://doi.org/10.1007/s10021-004-0008-z" target="_blank" rel="noopener">https://doi.org/10.1007/s10021-004-0008-z
久保田J. S.里格尔和V. A.拉扎尔,1970。阿拉斯加驼鹿食用牧草的矿物成分。野生动物学报34(3):565-569。<一个href="https://doi.org/10.2307/3798864" target="_blank" rel="noopener">https://doi.org/10.2307/3798864
刘,J, T.迪茨,S. R.卡朋特,M.阿尔伯蒂,C.福尔克,E.莫兰,A. N.佩尔,P.戴德曼,T.克拉兹,J. Lubchenco, E.奥斯特罗姆,Z.欧阳,W.普罗文彻,C. L.雷德曼,S. H.施耐德和W. W.泰勒。2007。人类与自然系统耦合的复杂性。科学》317(5844):1513 - 1516。<一个href="https://doi.org/10.1126/science.1144004" target="_blank" rel="noopener">https://doi.org/10.1126/science.1144004
马格努森,J·J, d·m·罗伯逊,b·J·本森,r·h·韦恩,d·m·利文斯通,t·阿莱,r·a·阿塞尔,r·g·巴里,v·卡德,e·库西托,n·g·格兰宁,t·d·普劳斯,k·m·斯图尔特,v·s·沃金斯基。2000。北半球湖泊和河流冰盖的历史趋势。科学》289(5485):1743 - 1746。<一个href="https://doi.org/10.1126/science.289.5485.1743" target="_blank" rel="noopener">https://doi.org/10.1126/science.289.5485.1743
麦克利兰,J. W., A.汤森-斯莫尔,R. M.霍姆斯,F.潘,M.斯蒂格利茨,M.科什和B. J.彼得森,2014。河流输出的营养物质和有机物从阿拉斯加北坡到波弗特海。水资源研究与开发,20(2):344 - 344。<一个href="https://doi.org/10.1002/2013WR014722" target="_blank" rel="noopener">https://doi.org/10.1002/2013WR014722
麦克道尔,G.福特,J.琼斯,2016。社区层面气候变化脆弱性研究:趋势beplay竞技、进展与未来方向环境科学研究11(3):033001。<一个href="https://doi.org/10.1088/1748-9326/11/3/033001" target="_blank" rel="noopener">https://doi.org/10.1088/1748-9326/11/3/033001
麦克拉伦,B. E.和R. O.彼得森,1994。皇家岛上有狼,驼鹿和年轮。科学》266(5190):1555 - 1558。<一个href="https://doi.org/10.1126/science.266.5190.1555" target="_blank" rel="noopener">https://doi.org/10.1126/science.266.5190.1555
梅雷迪思,M.萨默科恩,S.卡索塔,C.德克森,A.埃凯金,A.霍洛德,P.科菲纳斯,A.麦金托什,J.墨尔本-托马斯,M. M. C.米尔伯特,G.奥特森,H.普里查德和E. A. G.舒尔,2019。极地地区。h - o 203-320页。Pörtner, D. C.罗伯茨,V.马森-德尔莫特,P.翟,M.提格诺,E.波洛赞斯卡,K.明滕贝克,A. Alegría, M.尼科莱,A.奥肯,J.佩佐德,B.拉玛和N. M.韦耶,编辑。气候变化中的海洋和冰冻圈:政府间气候变化专门委员会(IPCC)的特别报告。beplay竞技IPCC,日内瓦,瑞士。
梅西耶,1994年。有蹄动物捕食种群模型:以北美驼鹿为例。生态75(2):478 - 488。<一个href="https://doi.org/10.2307/1939551" target="_blank" rel="noopener">https://doi.org/10.2307/1939551
莫尔德,1977年出版。阿拉斯加科尔维尔河地区驼鹿的运动模式。论文,阿拉斯加大学,费尔班克斯,阿拉斯加,美国。
Nakicenovic, N.和R. Swart,编辑。2000.排放场景。政府间气候变化专门委员会(IPCC)特别报告。beplay竞技剑桥大学出版社,英国剑桥。
尼莫,i.h., K.贝克曼,J. Godfroid, 2016。反布鲁氏菌驼鹿体内的抗体(Alces Alces gigas)、麝牛(Ovibos生于)和平原野牛(野牛野牛野牛野牛)在美国阿拉斯加。野生动物学报52(1):96-99。<一个href="https://doi.org/10.7589/2015-04-100" target="_blank" rel="noopener">https://doi.org/10.7589/2015-04-100
奥哈拉,t.m., G.卡罗尔,P.巴博萨,K.穆勒,J.布莱克,V.沃什纳和C.威莱托,2001。矿物和重金属含量与阿拉斯加驼鹿种群的死亡事件和招募不足有关。野生动物学报37(3):509-522。<一个href="https://doi.org/10.7589/0090-3558-37.3.509" target="_blank" rel="noopener">https://doi.org/10.7589/0090-3558-37.3.509
欧弗兰,j.e.,王敏,j.e.沃尔什,j.c.施特略夫,2014。未来北极气候变化:适应和减缓beplay竞技时间尺度。地球未来2(2):68-74。<一个href="https://doi.org/10.1002/2013EF000162" target="_blank" rel="noopener">https://doi.org/10.1002/2013EF000162
佩恩,C., S.潘达,A.普拉卡什,2018。阿拉斯加北坡Colville河上河流侵蚀的遥感。遥感10(3):397。<一个href="https://doi.org/10.3390/rs10030397" target="_blank" rel="noopener">https://doi.org/10.3390/rs10030397
彼得森,R. O.佩奇和K. M.道奇,1984。狼,驼鹿,以及种群周期的异速分布。科学》224(4655):1350 - 1352。<一个href="https://doi.org/10.1126/science.224.4655.1350" target="_blank" rel="noopener">https://doi.org/10.1126/science.224.4655.1350
Purtner H.-O。,D. C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, K. Mintenbeck, A. Alegriía, M. Nicolai, A. Okem, J. Petzold, B. Rama, and N. M. Weyer, editors. 2019. The ocean and cryosphere in a changing climate: a special report of the Intergovernmental Panel on Climate Change (IPCC). IPCC, Geneva, Switzerland.
罗林斯,M. A.蔡,S. L.施图弗,D.尼克斯基,2019。阿拉斯加北部流域径流和淡水输出的变化特征。冰冻圈13(12):3337-3352。<一个href="https://doi.org/10.5194/tc-13-3337-2019" target="_blank" rel="noopener">https://doi.org/10.5194/tc-13-3337-2019
阿拉斯加和北极规划情景网络(SNAP)。(无日期)。A2变暖情景下阿拉斯加降水的年代际摘要。SNAP,国际北极研究中心,阿拉斯加大学,费尔班克斯,阿拉斯加,美国。URL:<一个href="https://www.snap.uaf.edu/data" target="_blank" rel="noopener">https://www.snap.uaf.edu/data
西顿,C. T., T. F.帕拉吉,R. D.博尔杰,K.基兰,S.杜布瓦和C. L.弗莱纳,2011。浏览驼鹿生物量去除及营养状况酒精度酒精度.野生动物学报17(1):55-66。<一个href="https://doi.org/10.2981/10-010" target="_blank" rel="noopener">https://doi.org/10.2981/10-010
斯蒂芬妮,E., J.德拉格,D.米勒,B. M.琼斯,M.卡内夫斯基,2020。与通道迁移有关的塔里克斯,冷冻物,和永久冻土动力学,科尔维尔河三角洲,阿拉斯加。冻土与冰缘过程31(2):239-254。<一个href="https://doi.org/10.1002/ppp.2046" target="_blank" rel="noopener">https://doi.org/10.1002/ppp.2046
胶带,K. D.克里斯蒂,G.卡罗尔,J. A.奥唐纳。2016 a。北极的新野生动物:改变河岸生态系统和灌木栖息地扩张对雪鞋兔的影响。全球变化生物学22(1):208-219。<一个href="https://doi.org/10.1111/gcb.13058" target="_blank" rel="noopener">https://doi.org/10.1111/gcb.13058
胶带,K. D., D. D. Gustine, R. W. Ruess, L. G. Adams, J. A. Clark, 2016b。阿拉斯加北极地区驼鹿的活动范围扩大与变暖和增加的灌木栖息地有关。PLoS One 11(4): e0152636。<一个href="https://doi.org/10.1371/journal.pone.0152636" target="_blank" rel="noopener">https://doi.org/10.1371/journal.pone.0152636
胶带,K. D., M. Hallinger, J. M. Welker, R. W. Ruess, 2012。阿拉斯加北极地区灌木扩张的景观异质性。生态系统15(5):711 - 724。<一个href="https://doi.org/10.1007/s10021-012-9540-4" target="_blank" rel="noopener">https://doi.org/10.1007/s10021-012-9540-4
泰图斯,K. T. L.海恩斯,T. F.帕拉吉,2009。驼鹿、北美驯鹿、鹿和小型野味在阿拉斯加人饮食中的重要性。R. T. Watson, M. Fuller, M. Pokras和W. G. Hunt,编辑,第137-143页。从废弃弹药中摄入铅:对野生动物和人类的影响。游隼基金,博伊西,爱达荷州,美国。<一个href="https://doi.org/10.4080/ilsa.2009.0312" target="_blank" rel="noopener">https://doi.org/10.4080/ilsa.2009.0312
沃克,B.和D.索特,2006。弹性思维:在不断变化的世界中维持生态系统和人类。岛屿出版社,华盛顿特区,美国。
沃克,H. J.和P. F.哈德森,2003。阿拉斯加科尔维尔河三角洲的水文和地貌过程。地貌学56(3 - 4):291 - 303。<一个href="https://doi.org/10.1016/S0169-555X(03)00157-0" target="_blank" rel="noopener">https://doi.org/10.1016/s0169 - 555 x (03) 00157 - 0
Wam, H. K, A. M. Felton, C. Stolter, L. Nybakken, O. Hjeljord, 2018。驼鹿对桦树特定营养成分的选择限制了食物的可接受性。生态与进化8(2):1117-1130。<一个href="https://doi.org/10.1002/ece3.3715" target="_blank" rel="noopener">https://doi.org/10.1002/ece3.3715
韦斯科普夫,s.r., O. E.勒迪,L. M.汤普森,2019。beplay竞技气候变化对中西部鹿和驼鹿的影响。自然科学进展(4):769-781。<一个href="https://doi.org/10.1002/jwmg.21649" target="_blank" rel="noopener">https://doi.org/10.1002/jwmg.21649
温泽尔,g.w., 2000。努勒维特克莱德河的分享、金钱和现代因纽特人的生存义务和互惠。森里民族学研究53:61-85。
威伦斯基,1999。NetLogo(和NetLogo用户手册)。美国伊利诺斯州埃文斯顿西北大学互联学习与计算机建模中心。
r·J·沃尔夫和r·J·沃克,1987。阿拉斯加的自给经济:生产力、地理和发展影响。北极人类学24(2):56-81。
周杰,L. Prugh, K. D. Tape, G. Kofinas, K. Kielland, 2017。阿拉斯加北极地区植被结构在控制脊椎动物分布中的作用。地理科学进展,49(2):291-304。<一个href="https://doi.org/10.1657/AAAR0016-058" target="_blank" rel="noopener">https://doi.org/10.1657/AAAR0016-058
周杰,K. D. Tape, L. Prugh, G. Kofinas, G. Carroll, K. Kielland, 2020。北极地区增强的灌木生长增加了食草动物栖息地的连通性。全球变化生物学26(7):3809-3820。<一个href="https://doi.org/10.1111/gcb.15104" target="_blank" rel="noopener">https://doi.org/10.1111/gcb.15104
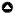
图1


图1所示。研究区域。红色表示纽克苏特驼鹿的活动区域,由猎人访谈和布劳德(2010)估算得出。绿色显示了可进入的狩猎区域,在这个场景中,河流的通航性增加了。黄线描绘了冰路的大致位置。虚线矩形是基于代理的模型中的模拟区域。乌米亚特是一个非建制社区,只有季节性居民,是该地区夏季研究活动和石油作业的营地和燃料站。
图2

图2所示。评估气候变化对驼鹿收获机会影响的社会-生态系统框架。beplay竞技
图3

图3。模拟了猎人数量、狩猎途径、驼鹿分布和丰度的变化。增加准入对驼鹿收获的影响有限。y轴为驼鹿收获量(kg)。x轴显示了四种情况,即当前(蓝色)和增加(橙色)河流通航通道。“H<年代ub>0米<年代ub>0是最近猎人数量(H)和驼鹿数量(M)的基线情景<年代ub>0米<年代ub>+的情况下,只有驼鹿的数量增加到历史最大值。“H<年代ub>+米<年代ub>0是猎人数量增加到历史最大值的情景。在" H "里<年代ub>+米<年代ub>+情况下,猎人数量和驼鹿数量都增加到历史最大值。H和M的增长百分比显示在柱状图的顶部。
图4

图4。驼鹿栖息地的扩张模式。绿色区域描述了IPCC A2在2020年20年代(A)、2050年(B)和2090年代(C)变暖情景下的驼鹿栖息地,采用Zhou et al.(2020)。蓝线代表驼鹿猎人可能使用的主要水道。红点表示纽克苏特和乌米亚特的位置。来自努伊克苏特的驼鹿猎人目前可以到达科尔维尔河上的乌米亚特,在其他主要流域也可以到达类似的距离。
图5

图5所示。研究区驼鹿预测。蓝线表示ARIMA模型估计的驼鹿平均数量。灰色和浅蓝色区域分别表示95%和80%的置信区间。
图6

图6所示。科尔维尔河在狩猎季节的流量和降水。(A) 2002-2020年(A1) 8月和(A2) 9月实测月平均降水量(Q)和估计月总降水量(P)的变化趋势。(B) 2002-2021年7月、8月和9月的平均Q和P与标准误差(误差条)。(C) PICC A2变暖情景下预测8、9月年代际平均降水量的标准误差(误差条)。问(m<年代up>3.年代<年代up>-1)在Umiat USGS站(https://www.usgs.gov/)测量,P (mm)在Umiat以上Colville河流域估计,基于SNAP数据(https://www.snap.uaf.edu)。
表1
表1。仿真模型中的场景矩阵。猎人和驼鹿的数量从目前(H<年代ub>0,米<年代ub>0)达到历史最大值(H<年代ub>+,米<年代ub>+)。
| 麋鹿数量 | |||
| 米<年代ub>0= 280 | 米<年代ub>+= 1535 | ||
| 猎人的数量 | H<年代ub>0= 12 | H<年代ub>0米<年代ub>0 | H<年代ub>0米<年代ub>+ |
| H<年代ub>+= 40 | H<年代ub>+米<年代ub>0 | H<年代ub>+米<年代ub>+ |
|
表2
表2。假设主要生存资源短缺的情景,以同等数量的驼鹿表示。“总流入”是进入Nuiqsut社区的所有收获资源的估计总量。“假设损失”是在假设的情况下,特定资源的短缺量,在“比例损失”中表示为总收获资源的百分比,在“驼鹿单位”中表示为等效的驼鹿数量。
| 一年 | 总流入(公斤) | 假设损失(kg) | 成比例的损失 | 驼鹿单位 |
| 假设鲸鱼数量全部减少 | ||||
| 1985 | 72591年 | 3383年 | 5% | 14 |
| 1993 | 121480年 | 34884年 | 29% | 143 |
| 2014 | 168733年 | 67171年 | 40% | 275 |
| 场景2假设驯鹿全部损失 | ||||
| 1985 | 72591年 | 27225年 | 38% | 112 |
| 1993 | 121480年 | 37271年 | 31% | 153 |
| 2014 | 168733年 | 47715年 | 28% | 196 |
| 假设三文鱼全部损失(斑鳟属撒拉族) | ||||
| 1985 | 72591年 | 620 | 0.9% | 3. |
| 1993 | 121480年 | 458 | 0.4% | 2 |
| 2014 | 168733年 | 1764年 | 1.0% | 7 |
| 假设非鲑鱼鱼全部损失 | ||||
| 1985 | 72591年 | 31408年 | 43% | 129 |
| 1993 | 121480年 | 40588年 | 33% | 166 |
| 2014 | 168733年 | 38604年 | 23% | 158 |
| 场景5假设密封圈全部损失 | ||||
| 1985 | 72591年 | 2010年 | 3% | 8 |
| 1993 | 121480年 | 3769年 | 3% | 15 |
| 2014 | 168733年 | 9652年 | 6% | 40 |
| 假设鸟类和鸡蛋全部损失 | ||||
| 1985 | 72591年 | 3645年 | 5% | 15 |
| 1993 | 121480年 | 1962年 | 2% | 8 |
| 2014 | 168733年 | 2203年 | 1% | 9 |