






以下是引用这篇文章的既定格式:
先生,先生,p.o 'B。莱弗,A. M.戈姆利,N. J.斯科特,A. R.麦金托什,D.弗莱彻,J.提利亚纳基斯,2022。一个人口统计模型,以支持习惯管理一个文化上重要的水禽种。生态与社会27(3):14。摘要
土著人民和当地社区(IPLC)为协助习惯环境管理而邀请的科学支持可以改善保护和社区生计。例如,人口模型可以帮助了解备选野生动物管理战略如何影响种群动态和收获可持续性。我们开发了一种人口统计模型,以协助新西兰Aotearoa最南端的毛ori部落ngongi Tahu对文化上重要的黑天鹅族群(谷湖īanau,天鹅座atratus会).我们使用了最近的人口统计数据,包括卵子收获实验研究的结果新西兰tiaki(ngongi Tahu环保卫士),讨论了习惯的采蛋和来自1岁以上天鹅运动狩猎的背景压力对种群增长的差异影响。我们还评估了天鹅狩猎运动如何影响传统蛋的可持续性。估计人口增长(1.018或目前每年增长1.8%;95% CI: 0.808 ~ 1.241)对成虫和亚成虫存活率变化最敏感,其次为幼虫(1年)存活率、繁殖倾向和巢孵化成功。种群增长的不确定性几乎完全归因于天鹅孵化后存活率的不确定性。根据成虫和亚成虫存活率的微小变化,可持续种群水平的卵子收获率从零到过半不等。种群对成虫和亚成虫生存的敏感性表明,限制和监测它们的死亡率对种群和卵收获的可持续性至关重要,而当代政府通过新西兰渔业和狩猎实施的物种管理,允许运动狩猎造成成虫和亚成虫的死亡率,几乎没有收获的记录。认识到ngongi Tahu的权利和利益,更密切地监测天鹅的死亡率,可以提高ngongi Tahu实行习惯收割的能力,加强人口和环境监测,并在适当时以文化上适当和较少浪费的方式控制天鹅数量。我们提出的模型可以在未来潜在的共同管理安排中,帮助ngangui Tahu和新西兰政府之间的决策和沟通。人口模型可以成为支持习惯环境管理的有用工具,但开发、维护和实施这些工具需要支持具有适应性的政策和管理安排,这些政策和管理安排承认IPLC对环境和决策的权利。介绍
当代环境管理系统正在努力遏制生物多样性和生态系统健康的全球下降(Díaz等,2019年,IPBES 2019a)。管理的一个限制因素是在大范围内以高分辨率持续监测生物多样性和生态系统的能力,以及在当地实施适应性保护措施的能力(Cumming等,2006年,Guerrero等,2013年)。为了帮助克服这一限制,并更普遍地要求对环境的权利和责任,许多土著人民和当地社区(IPLC或<年代pan class="ob-q">”与最近定居、占领或殖民某一地区的族群相比,是某一地区原始居民的后裔和认同的族群<年代pan class="ob-q">”;IPBES 2019b)在授权其习惯环境管理系统方面积极发言(Ens等人,2012年;Witter等人,2015年;Lyver和Tylianakis, 2017年)。在许多习惯的管理系统中,IPLC不断地与环境进行接触和监测,同时随着时间的推移,以精确的分辨率积累生物多样性和生态系统的长期经验知识(Gadgil等人1993年,Moller等人2004年,Berkes 2009年)。习惯上的资源使用和与环境的接触支撑着IPLC福祉的许多方面,包括生计、文化认同和社区亲属关系,而这些关系可以激励可持续的实践和地方保护的执行(Cinner和Aswani, 2007年,Lyver等人,2008年,Halim等人,2012年,Sheil等人,2015年,Caillon等人,2017年)。因此,支持IPLC对其土地和资源进行常规管理可能是加强环境监测和保护的一种实用和公正的方式(Sobrevila 2008年,Artelle等人2019年,Herse等人2020年,Dawson等人2021年,Reyes-García等人2022年)。
在许多地方,IPLC在决定是否可以使用资源、什么用途和策略是合适的以及应该允许谁使用方面的法律权威要么缺失,要么被忽视(Kinan和Dalzell 2005年,Goldman 2011年,ruu等人2017年,Snook等人2020年)。在这种情况下,非IPLC当局的管理决策往往倾向于娱乐或商业收获的竞争利益(Ban等人,2017年)、其他土地用途,如采矿(Parlee等人,2018年),或保护主义策略,如保护区,优先考虑IPLC与其环境之间的分离(Brockington和Igoe, 2006年,Solomon 2014年),不重视习惯收获(Hunn等人,2003年,Tyrrell 2008年,Lyver等人,2019a)。环境科学家越来越多地使用动物种群的人口模型来为习惯资源使用和管理的决策提供信息(Brook et al. 2006, Zador et al. 2006, Bradshaw and Brook 2007, Chaloupka and Balazs 2007, Jones et al. 2015, McKechnie et al. 2020)。例如,Chaloupka和Balazs(2007)使用人口统计模型为美国夏威夷的公共政策辩论提供信息,讨论重新开始绿海龟的习惯捕捞的可持续性(龟鳖目mydas),在商业捕捞过度开采及随后将绿海龟列为濒危物种后,这些物种被禁止<年代pan class="ob-q">”濒临灭绝的物种。<年代pan class="ob-q">”在澳大利亚,Bradshaw和Brook(2007)使用人口模型来评估平衡外来濒危banteng习惯使用的策略。Bos javanicus),并承担娱乐和商业收获。然而,如果不是由IPLC发起或不包括IPLC代表,IPLC对科学证据的声音和解释在此类研究中很大程度上是缺席的。同样,当IPLC希望参与习惯管理并欢迎科学支持时,与IPLC合作收集、解释和传播数据对于确保其权利、价值和优先事项得到充分承认非常重要(Ens等人,2012年;Rayne等人,2020年;Dawson等人,2021年)。
在这里,我们提出了一个人口统计模型,该模型是与新西兰Aotearoa地区最南端的毛ori部落nongi Tahu合作开发的,以支持黑天鹅的习惯管理(谷湖īanau,天鹅座atratus会)在一个200平方公里的沿海泻湖和关键的黑天鹅繁殖区,Te Waihora/埃尔斯米尔湖(图1)。黑天鹅及其卵的收获一直是Te Waihora世世代代的习惯管理和食物采集的重要组成部分,但自新西兰殖民政府在公元20世纪初控制了当地物种管理以来受到限制(Lamb 1964年,Pauling和Arnold 2008年,Herse等人2021年)。新西兰现行法律将黑天鹅归为<年代pan class="ob-q">”gamebirds<年代pan class="ob-q">”可由猎鸟许可证持有人狩猎作为运动(新西兰政府1953年),但禁止未经特别许可的惯例收割(Herse等人,2020年,2021年)。当代政府授权的野鸟管理人员使用每年设定的每日袋数限制(配额)来维持天鹅的数量,以满足运动猎人的娱乐利益(新西兰政府1987年),过去还允许当地农民(欧洲人新西兰人)捕杀,以限制放牧和潜在的经济损失(Williams 1979年)。由于当代的猎禽管理机构新西兰渔业与狩猎(FGNZ)不要求有执照的狩猎者或土地所有者报告其猎捕趋势,因此人们对运动狩猎和过去扑杀的压力程度知之甚少,FGNZ对猎捕趋势的定期区域电话调查也不针对特定地点(Herse等人,2020年)。
ngongi Tahu对管理天鹅数量感兴趣,至少在一定程度上是通过未经监督或特别许可的自行授权的习惯采卵(Herse等人,2020年,2021年)。ngongi Tahu自主授权收割和管理天鹅的权利在新西兰的建国文件中得到了保障新西兰(这片土地的原居民)和英国政府,1840年签订了怀唐伊条约<年代pan class="ob-q">”英格兰女王同意保护新西兰的酋长、亚部落和所有人民,不受限制地行使他们对土地、村庄和所有财富的酋长统治权<年代pan class="ob-q">”(Kawharu 1989:321)。同样的,ngangui Tahu的权利<年代pan class="ob-q">”确定和制定开发或使用其土地或领土和其他资源的优先事项和战略<年代pan class="ob-q">”受到《联合国土著人民权利宣言》(联合国大会2007:23)的保护。然而,在现行的野鸟管理法规下,这些权利尚未实现或颁布。除了fgnz允许的运动狩猎和扑杀,1968年的一场大风暴对水生植物造成的破坏<年代pan class="ob-q">”太太<年代pan class="ob-q">”风暴),再加上过去半个世纪大量的农业径流,已经使Te Waihora退化,耗尽了天鹅的大型植物食物供应(Lineham 1983, Gerbeaux 1989, Hearnshaw and huhey 2010),天鹅数量减少了约90% (Williams 1979, Ford et al. 2017)。因此,尚不清楚目前的天鹅数量是否能够维持潜在的习惯采卵或任何正在进行的狩猎运动,以及天鹅的生育能力(孵化成功)和生存压力分别如何影响种群数量。
我们的目标是向ngangui Tahu提供关于Te Waihora天鹅数量的最新信息,以便与FGNZ和新西兰政府讨论重新开始传统采蛋的潜在策略。为此,我们首先使用最近的人口数据和一个年龄分类的矩阵人口模型(a)估计当前的人口年增长率或人口规模的变化(λ;卡斯韦尔2001)。然后,我们使用该模型来评估(b)天鹅生育率和年龄特异性存活率的变化如何不同地影响λ,以及(c)这些人口统计率的当前不确定性水平如何不同地促进λ的不确定性。为了理解运动狩猎的卵收获等价性,我们估计了(d)新鲜下蛋和孵化蛋的预期终身产量(即生殖价值的度量;Caswell 2001),这些天鹅的年龄≥1岁,集体最容易被射杀(Williams 1977)。最后,我们使用了最近一项卵子采集实验研究的结果(Herse等人,2021年)来(e)评估体育狩猎对ngongi Tahu潜在的习惯卵子采集的可持续性的影响。基于模型的结果,我们探索了将习惯的卵采集纳入当地物种管理的潜在途径。最后,我们概述了改善对习惯管理的支持的潜在方法,这可以加强环境监测和保护(Ens等人,2016年,Herse等人,2020年,Dawson等人,2021年,Lyver等人,2021年)。
方法
研究物种和种群
现代的黑天鹅(c . atratus会)源自澳洲,与当地特有的新西兰黑天鹅(阿宝ū佤邦,c . sumnerensis),大约在15世纪,在毛ori人到达新西兰之后,在欧洲人到达之前,它在新西兰大陆上灭绝了(罗伦斯等人,2017年)。欧洲人报道c . atratus会19世纪在新西兰是没有的,19世纪60年代从澳大利亚引进了几对(兰姆1964)。然而,随着物种的引入,人口的快速增长,以及最近的DNA证据,表明c . atratus会在19世纪之前和期间从澳大利亚传播,和/或通过与引入的个体混合对现存种群的基因拯救,可能已经发生过(Rawlence等,2017,Montano等,2018a)。因此,科学家们一直在争论现代黑天鹅是否原属于新西兰(Montano等,2018b, Rawlence等,2018)。一些ngaoi Tahi部落成员认为黑天鹅原属于新西兰(Bataille 2020),在ngaoi Tahu和自然保护部长之间的Te Waihora联合管理计划(Te rongnanga o ngaoi Tahu和自然保护部2005年)中,该物种被确认为黑天鹅。更重要的是,黑天鹅被一些恩加西塔胡人视为文化实践的关键元素,比如kaitiakitanga(环境监护)mahinga凯(<年代pan class="ob-q">”习惯上的食物和自然材料的采集以及这些资源的采集地<年代pan class="ob-q">”;新西兰政府1998:102,Bataille 2020, Bataille等人。2021),这对马霍里文化完整性(kawallu 2000)、知识生产(Lyver等人。2019b)和保护(Wehi和Lord 2017)很重要。该物种也受到一些野鸟许可证持有者和其他公众的重视,而一些农民认为天鹅是害虫,因为它们有时在牧场上吃草(Bataille等人,2020,2021)。
“黑天鹅”(以下<年代pan class="ob-q">”天鹅<年代pan class="ob-q">”)主要在浅湖、泻湖和河口繁殖(Williams 1981, 2013a)。Te Waihora拥有新西兰南岛最大的天鹅种群,在过去20年里,估计其种群规模从5000到10000只不等(Ford et al. 2017, Herse et al. 2020)。与全球其他天鹅物种相似,Te Waihora的大多数黑天鹅寿命不到10年,每年只有不到一半的成年黑天鹅尝试筑巢(Williams 1979, 1981, 2013, Bart et al. 1991)。黑天鹅通常是一夫一妻制的,在特怀霍拉岛的繁殖通常是同步的,从8月到9月,随着冬季水位下降,在大多数巢穴孵化后的10月开始减少(Williams 1981, 2013a, Herse et al. 2021)。平均窝卵大小为5-6颗(Williams 2013a, Herse等人,2021)。第二轮繁殖有时会在10月至12月期间进行,其中包括在给定的繁殖季节中约三分之一的筑巢(唐纳德·布朗,ngongi Tahu,新西兰基督城,2018年6月19日,个人沟通;Herse等人,2021年)。在之前的一项实验卵采集研究中,Herse等人(2021年)发现,在完全去除第一批卵后,天鹅倾向于替换一部分这些卵,这样,在收获的巢穴中,平均巢级孵化成功为2.61只小天鹅(cf.每未收获的巢穴5.68只小天鹅)。尽管Herse等人(2021年)无法确定离合失败后重新筑巢是由原巢居住者还是其他使用新巢的天鹅进行的,但已经有个别天鹅对在离合失败后重新筑巢的记录(Williams 1979年),这表明至少有一些第二轮繁殖是由于重新筑巢。在我们的研究区域,对鸟巢孵化成功的压力包括巢穴破坏、洪水和不育,但在最大的筑巢群体中,孵化成功通常很高(Adams 1971, Meis 1988, Herse等人,2021)。幼天鹅(小天鹅)在孵化后约3-5个月达到独立,在2-4岁时性成熟(Williams 2013a)。
除了食物供应,目前天鹅生存的压力还包括体育狩猎(Williams 1979, Herse et al. 2020)。Gamebird管理人员分别根据定期航空调查和电话调查估计的天鹅数量趋势和承载量设定袋限,并在适用袋限的狩猎区域或分区域范围内进行汇总(Herse等人,2020年)。近年来,在包括Te Waihora在内的约2500平方公里狩猎分区域内,猎鸟许可证持有者在5月至7月的大部分时间内每人每天可射杀5只天鹅,在8月至9月的大部分时间内每人每天可射杀20只天鹅(Herse等人,2020年)。然而,由于几个原因,人们对Te Waihora野鸟目前的射击率及其对天鹅数量的影响知之甚少。新西兰法律要求私人土地所有者遵守当地猎鸟袋的限制,但不要求土地所有者在他们的财产上射击天鹅获得许可证(新西兰政府1953年),或报告他们的猎取。同样,FGNZ不要求猎禽许可证持有者报告他们的起飞。此外,区域采油调查并不针对具体地点,而是包括特怀霍拉分区域以外的地区(Herse等人,2020年)。
矩阵人口模型
我们建立了一个年龄分类的矩阵种群模型(Leslie 1945, Caswell 2001)来描述天鹅的种群动态。该模型定义为
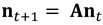 |
(1) |
在哪里<年代trong>一个预测矩阵是否包括特定年龄、人均天鹅生育率和存活率的估计,以及<年代trong>nt是一段时间内每个年龄组的个体数量的向量吗t(卡斯韦尔2001)。时间单位为一年,从蛋孵化时开始,到下一个产卵季节孵化前结束(<年代pan class="ob-q">”post-breeding普查<年代pan class="ob-q">”;卡斯韦尔2001)。我们假设该种群对移民和移民是封闭的,因为天鹅在该区域内的水体中很少分散(Williams 1977;赫斯,莱弗,斯科特,提利亚纳基斯和恩加西·塔胡,未发表的数据),本地物种管理无法控制种群中少数分散个体的移动。
考虑到第一次繁殖的年龄为2-4岁(Williams 2013a),根据孵化、性不成熟亚成虫和成虫阶段分类建立一个阶段分类模型是合理的。但是,由于在人口普查中无法区分亚成虫和非繁殖成虫,我们无法估计每年尝试繁殖成虫的比例;我们只能估计亚成人期和成人期的这个比例。此外,我们缺乏关于Te Waihora天鹅当前年龄分布的数据,也不知道运动狩猎的持续死亡率是否或如何在不同年龄层之间变化。因此,我们基于一个简化的生命周期,包含孵化年(HY,年龄<孵化1年)和孵化年后(AHY,年龄≥孵化1年)年龄分类(图2)。该模型中的生育期包括每年尝试繁殖的AHY天鹅的比例,即亚成体加成体,我们可以估算。更精细的年龄或阶段类别的人口比例可以在以后使用附加数据参数化,如果这些数据可用的话。
该模型可以写成矩阵形式为
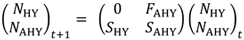 |
(2) |
在哪里NHY,t而且NAHY,t分别为时间(年)内HY和AHY天鹅的数量,t;FAHY为年繁殖力或每只AHY天鹅孵化的卵数;年代沪元天鹅在孵化后第一年存活的概率是多少<年代pan class="ob-q">”孵化一年<年代pan class="ob-q">”);而且年代AHY为AHY天鹅的年生存概率。HY天鹅的繁殖力(公式2中矩阵左上角的元素)为0,因为HY天鹅不繁殖(Williams 2013)。建模的人口转换率(矩阵元素)是时不变的,但可以每年用新数据更新。
影响AHY天鹅年繁殖力的因素包括尝试繁殖的倾向、鸟巢孵化的成功和AHY天鹅的死亡率(Williams 1979, 1981, Herse et al. 2021)。未收获和收获的鸟巢的孵化成功不同,因为近年来,天鹅往往只替换收获的蛋的一个子集(Herse等人,2021年)。因此,我们估计FAHY使用表达式:
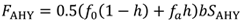 |
(3) |
在哪里f0每个未收割的巢的生育能力,f一个每收获一个巢的生育能力,h总巢收获的比例,和b为试图繁殖的AHY天鹅的比例,如上所述。每个巢的生育能力乘以0.5就等于每只繁殖天鹅的生育能力,因为繁殖通常是一夫一妻制的(Williams 2013a)。该模型假设,在固定产量的鸡蛋收获过程中,收获者从收获的巢穴中移除整批鸡蛋(而不是从数量更多的巢穴中移除更少的鸡蛋),因为在最近的收获实验中发现,这是对种群水平孵化成功影响最小的策略(Herse等人,2021年)。的年代AHY公式3中的项说明了有些天鹅在当年没有存活下来,打算在来年繁殖t+ 1。
在Te Waihora,大型植物食物的有限可获得性可能会阻止天鹅数量增加到一定规模以上,不管来自体育狩猎或习惯收获的压力(Williams 1979, Herse等人,2021)。然而,我们没有在模型中包含密度依赖性,因为我们缺乏定义Te Waihora天鹅种群承载能力的信息。
人口统计参数的估计
我们使用包括经验数据和间接方法的参数自举方法,分三步估计人口统计参数和相关方差(图3)。首先,我们使用最近的人口统计数据来定义λ,年代AHY的基础参数FAHY(方程3,表1,图3)。其次,我们从初始分布中使用随机抽样进行反计算,从而生成一个bootstrap分布的估计值年代沪元,我们缺乏数据来自信地直接估计(表1,图3)年代沪元合并了关于天鹅幼仔生存的信息,结果调整了bootstrap分布年代AHY和潜在的参数FAHY(表1,图3)。最后,我们从得到的bootstrap分布中使用随机抽样来参数化矩阵模型,并为Te Waihora种群的一组人口参数生成bootstrap分布(表2,图3)n= 10000个样本。我们将在下面和附录1中详细介绍这些步骤。
定义用于回算的初始抽样分布年代沪元,我们估计N沪元而且NAHY根据两名独立观察员在2018年12月繁殖后空中普查期间的天鹅平均数量(图3,见附录1)f0而且f一个使用的数据分别来自2018年筑巢高峰期(8 - 9月)在Te Waihora进行的一次实验采蛋过程中对未采蛋和已采蛋巢穴的观察(图3、图A1.1,见附录1;Herse等人,2021年)。因此,该模型假设某一年的所有繁殖都发生在筑巢高峰期,我们直观地估计这包括2018年Te Waihora所有巢穴的约88%(见附录1)。然而,该模型考虑了季末筑巢对估计种群增长率的贡献,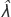 ,间接通过反计算年代沪元(详见下文),其中包括关于N沪元以及整个种群的大小,包括从晚季巢穴孵出的小天鹅。我们估计b为在2018年Te Waihora筑巢高峰期试图繁殖的AHY天鹅的比例,由巢穴的空中普查确定(假设每个巢穴代表两只繁殖天鹅;Williams 2013a)和AHY天鹅(图3,图A1.2,见附录1)b2018年(0.30)与之前研究的估计相似(0.16-0.34;威廉姆斯1981)。我们估计年代AHY基于2019-2020年期间我们在Te Waihora监测的17只gps标记的AHY天鹅的已知命运(图3,见附录1),其估计的平均生存率与之前长期带状恢复研究的估计相似(图3,图A1.3;威廉姆斯1979年,巴克和布坎南1993年)。
,间接通过反计算年代沪元(详见下文),其中包括关于N沪元以及整个种群的大小,包括从晚季巢穴孵出的小天鹅。我们估计b为在2018年Te Waihora筑巢高峰期试图繁殖的AHY天鹅的比例,由巢穴的空中普查确定(假设每个巢穴代表两只繁殖天鹅;Williams 2013a)和AHY天鹅(图3,图A1.2,见附录1)b2018年(0.30)与之前研究的估计相似(0.16-0.34;威廉姆斯1981)。我们估计年代AHY基于2019-2020年期间我们在Te Waihora监测的17只gps标记的AHY天鹅的已知命运(图3,见附录1),其估计的平均生存率与之前长期带状恢复研究的估计相似(图3,图A1.3;威廉姆斯1979年,巴克和布坎南1993年)。
我们使用上述初始引导分布中的随机样本进行反向计算年代沪元.因为人口增长率,λ,是矩阵的优势特征值<年代trong>一个,可由特征方程求解:
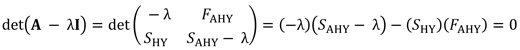 |
(4) |
在哪里<年代trong>我是一个2 × 2单位矩阵(Caswell 2001),我们重新排列公式4,并估计年代沪元使用表达式:
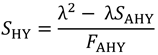 |
(5) |
其中λ是根据2017-2019年Te Waihora天鹅种群规模的年度趋势估算的,由航空普查确定(见附录1)。具体而言,我们估计公式5中的λ是对数种群规模与年份相关的线性回归曲线斜率的指数(图3、图A1.4;见附件1;Peery等人。2006)。我们对方程5应用了几个约束条件以确保<年代pan style="font-family:Symbola; font-size: 115%">Ŝ沪元>是否为0且小于患者的估计生存率n= 2018年,在Te Waihora出生的头40天里,有50只标记的天鹅幼崽(图3;如果在抽取样本的过程中没有满足约束条件,我们将丢弃该抽取的所有参数的样本,并重新采样。的结果引导分布的平均值年代沪元(表1、图3)与根据天鹅孵化数据的生存分析和以往研究(图A1.5,见附录1;威廉姆斯1979,梅斯1988)。约束条件在反计算中的应用年代沪元结果调整了反向计算中使用的参数的bootstrap分布(表2,图3)。
我们使用上述自举分布中的随机样本来参数化矩阵模型(表1、图3)<年代pan class="ob-q">”popbio<年代pan class="ob-q">”软件包(Stubben et al. 2020, R Core Team 2021)来估计天鹅种群的一套人口统计参数,假设没有采卵压力(h= 0)。我们估计年度λ作为矩阵的主导特征值<年代trong>一个(图3),如上所述(Caswell 2001)。以确定如何变化年代沪元而且年代AHY,以及较低水平的FAHY(f0而且b),差异影响λ,我们计算灵敏度(年代<年代ub>x)和弹性(e<年代ub>x)以绝对和比例的变化,分别在每个潜在的人口比率x(见附件1;卡斯韦尔2001)。弹性可以解释为一个给定的人口比率在决定λ方面的相对重要性的度量(Caswell 2001)。敏感性和弹性都是有用的,它们只是衡量不同尺度(即绝对比例)的人口比率变化的影响;对两者的测量确保了我们不会忽视只在一个尺度上明显的影响(卡斯韦尔2001年)。为了评估当前人口比率的不确定性水平如何不同地贡献了λ的不确定性,我们使用了的自举分布的方差年代<年代ub>x而且e<年代ub>x计算λ中每个人口统计率对总方差的比例贡献(见附录1;卡斯韦尔2001)。为了确定从种群中去除一只AHY天鹅的卵收获当量,我们估计了预期寿命和生殖价值(即人均一生的预期生殖产量,r沪元,或产生HY天鹅的数目;Caswell 2001),然后估计了预期的人均终身产卵的鲜蛋数
 |
(6) |
在哪里年代蛋是未收获巢穴中单个蛋的估计平均存活(孵化)概率(Herse等人,2021年)。
混合经营的可持续性
为了评估AHY天鹅的运动狩猎如何影响潜在的习惯卵收获的可持续性,我们使用矩阵模型估计λ基于的不同比率年代AHY而且h单独或结合。正如上文所述(Herse et al. 2020),特怀霍拉特有的天鹅运动狩猎信息并不存在年代AHY解释了这种压力带来的风险。因此,我们考虑了一系列年代AHY覆盖模型估计平均值年代AHY±0.1(以0.02为增量),这是一个足够大的范围来检测阳性和阴性在所有可能的速率中h.我们考虑了所有的潜力h(0-1,以0.1为增量),因为在20世纪早期和中期,天鹅种群曾维持着较高的脱卵率(Lamb 1964, Williams 1979)。对于每一个年代AHY而且h,我们保持这两个速率恒定,并使用n=从上面描述的其他参数的bootstrap分布的每个随机样本中得到10000个λ。这种方法允许我们评估不确定性对于每一个年代AHY而且h.
结果
我们用于天鹅种群动态建模的人口统计数据与以往研究的数据一致(表1;见附录1),该模型提供了生态上合理的人口统计参数估计(图3、4、表2)。
人口比率对人口增长的差异影响
λ的自举分布均值为1.018(1.8%的年增长率)(95% CI 0.808-1.241)。灵敏度和弹性分析都表明,λ受到的影响最大年代AHY的变化年代沪元,b,最不重要的是,f0(表2,图3)。基于的bootstrap分布的均值年代<年代ub>x而且e<年代ub>x的绝对变化的影响年代AHY的λ约为的30倍f0,而成比例变化的影响年代AHYλ几乎是λ的六倍f0(表2)。λ的不确定性几乎完全是由于年代沪元而且年代AHY(表2)。
孵化后天鹅的预期寿命和繁殖价值
该模型估计AHY天鹅的预期寿命为5.7年(95% CI: 3.4-17.0;(见附录1),假设无采卵压力,每只AHY天鹅一生中产生的平均鲜卵数和孵化卵数分别为5.9 (95% CI: 2.1-20.2)和4.5 (95% CI: 1.7-15.3)(图3)。
混合经营的可持续性
的bootstrap分布的均值年代AHY,该模型估计,在Te Waihora所有天鹅巢中,能够可持续地收获所有第一批孵化的蛋的比例约为0.3(图4)。然而,该种群能够维持的蛋的收获水平很大程度上取决于年代AHY(图4)。例如,如果……,所有天鹅巢中可持续收获第一窝卵的估计比例将降至零年代AHY只降低0.02,大约翻倍到~0.6年代AHY增加0.02(图4)年代AHY增加了0.04,该模型估计从Te Waihora的第一批鸡蛋中收获几乎所有的鸡蛋是可持续的(图4)。
讨论
可持续的物种管理需要了解种群如何应对不同的人口压力,并不断调整管理以适应种群和环境波动(Moller et al. 2004, Weinbaum et al. 2013)。Te Waihora天鹅种群对亚成体和成体(AHY个体)的生存变化比生育能力的变化更敏感,这与其他长寿水禽物种一致(Cooch et al. 2014, Koons et al. 2014, Wood et al. 2019)。此外,种群对生育能力的敏感性主要是由于对繁殖倾向的潜在敏感性,而不是对巢孵化成功的敏感性。此外,目前天鹅种群增长率的不确定性几乎完全归因于天鹅孵化后存活率的不确定性(表2)。这些结果可能特别有助于为加强种群监测和收获管理的潜在努力提供信息。
我们的模型的局限性反映了与收集动物种群人口统计数据相关的一些挑战。具体来说,HY天鹅生存的间接估计,以及估计AHY天鹅生存的样本量小,导致模型估计的不确定性很大。对长寿物种存活的更可靠的估计是基于长期监测(例如,标记-再捕获)项目的数据,其中包括数百或数千次观察(Sandercock 2006)。这种监测项目的开发和维护成本很高,在新西兰已经有几十年没有对天鹅进行过监测了(Williams 1977, 1979, Barker and Buchanan 1993)。尽管如此,我们对天鹅存活率的估计与之前基于长期标记-再捕获数据集对新西兰天鹅的估计一致(图A1.3,图A1.5;Williams 1977, 1979, Barker and Buchanan 1993),以及对全球其他天鹅物种的估计(Bart et al. 1991),表明它们至少是现实的。然而,在我们的模型中缺乏关于密度依赖性的信息,可能导致在一些建模管理场景下,对种群增长的估计过高(图4)。例如,AHY的生存和种群增长可能至少部分受到大型植物食物的可获得性的调节,而Te Waihora地区的植物食物是稀缺的(Lineham 1983, Gerbeaux 1989, Ford等人2017),这可能会阻止AHY的生存增加到我们考虑的一些水平。尽管狩猎活动可能减少。更可靠的数据(例如,来自长期监测项目的数据)可以提高精确度,减少我们估计的潜在偏差,并允许纳入额外的人口统计学参数,如密度依赖性。此外,更精细的年龄或阶段分类数据有助于解释潜在的瞬态种群动态(Koons等人,2005年,Ezard等人,2010年),我们发现这是次要的(图A1.6)。 Despite these data limitations, our model was useful for assessing differential effects of age-specific demographic rates on population growth, and how offtake from sport hunting affects the sustainability of potential customary egg harvests by Ngāi Tahu.
管理的影响
我们的研究结果引起了人们对FGNZ当代物种管理的关注,这对ngangui Tahu的习惯管理有一定的影响。Te Waihora天鹅的数量对AHY天鹅的死亡率高度敏感,但FGNZ通过允许的(大部分)AHY天鹅的运动狩猎来管理天鹅数量,通过固定的配额进行管理。即使是对种群维持体育狩猎能力的轻微高估,如果不通过增加存活和/或繁殖来补偿,也可能导致种群数量下降(Pöysä et al. 2004, Sandercock et al. 2011),并阻碍可持续的传统卵子收获。由于天鹅狩猎季节(5月至9月)与繁殖季节(8月至9月筑巢高峰)重叠,特怀霍拉岛周围农场的猎人群体有时会在天鹅准备筑巢时射杀大量天鹅(> 100)(唐纳德·布朗,新西兰基督城ngwi Tahu, 2021年2月9日,个人沟通).这种活动可能会对种群产生不成比例的巨大影响,不仅减少了每年的成虫存活率,还减少了试图繁殖的成虫的比例,特别是如果在筑巢前的射击破坏了促进同步殖民地繁殖的社会线索(Burger 1988, Reed和Dobson 1993, Wilhelm和Storey 2002)。新西兰tiaki和传统的采蛋者报告称,在特韦霍拉岛最大的天鹅筑巢区,繁殖的天鹅数量下降了,这至少部分可能是由于天鹅在筑巢前或筑巢期间立即被杀死(唐纳德·布朗和克雷格·鲍林,新西兰克莱斯特彻奇ngongi Tahu, 2021年2月9日,个人沟通).从本质上说,土地所有者正在使用FGNZ野鸟法规来控制他们所在地区的天鹅数量,以限制天鹅对新草和作物的潜在影响。此外,如上文所述,FGNZ不监测天鹅的基线死亡率,也不要求土地所有者或猎鸟许可证持有人报告其猎鸟量。考虑到本研究和相关研究(Herse等人。2020年和2021年),以及当地人表达的担忧新西兰tiaki最近,该地区FGNZ的管理人员表示,他们计划调整当地的野鸟法规,以减轻特怀霍拉天鹅的运动狩猎压力。然而,在FGNZ管理中缺乏监测和由此产生的不确定性限制了我们的人口模型的效用,以及能力新西兰tiaki评估在当前报告环境下的可持续卵子收获率。因此,有必要更密切地监测天鹅存活率的基线率和狩猎的影响,以确保配禽法规是可持续的,不妨碍可持续的习惯收获,这是本研究提出的一个关键建议。
充分承认ngangui Tahu的权利和利益,并赋予权力新西兰tiaki在当地物种管理方面的权威,可以带来双赢的结果,改善人口和环境监测(Herse等人,2020年),并在适当时控制天鹅数量,而不浪费资源或不必要地杀死天鹅(Bataille 2020年,Bataille等人,2021年,Herse等人,2021年)。新西兰tiaki和传统的采卵机可以通过不断生成关于繁殖活动(如筑巢群体的位置和大小、孵化成功)和环境压力(如水污染、洪水、巢穴破坏)的信息,加强对天鹅和特韦霍拉岛的监测,而这些信息是FGNZ目前在当地物种管理中所缺乏的(Herse等人,2020年和2021年)。为了控制天鹅数量,限制问题地区牧场的放牧,习惯收获也可能是一种实用的、文化上合适的、浪费较少的替代选择(Bataille 2020年,Bataille等人2021年,Herse等人2021年)。新西兰tiaki有权决定维持何种水平的天鹅数量是合适的(联合国大会2007年,橙色2011年),而目前的决策偏向于运动猎人和农民的利益(新西兰政府1987年,Bataille 2020年,Bataille等人2021年)。尽管特怀霍拉岛周围的大多数土地所有者重视湿地健康,但活跃的农民通常优先考虑经济收益,这促使一些人射击天鹅,以限制潜在的放牧影响(Bataille 2020, Bataille等人2021)。具有讽刺意味的是,天鹅放牧牧场可以部分归因于农业污染造成的大型植物稀缺(Lineham 1983, Gerbeaux 1989, Hearnshaw and huhey 2010, Williams 2013a)。因此,在采取当地天鹅数量控制措施之前,减少天鹅放牧对农民的影响应该优先控制污染,污染会使Te Waihora退化,导致天鹅在牧场上觅食(Williams 1979, 2013a)。然而,如果新西兰如果国王能做出一个公平的决定来控制天鹅的数量,那么习惯的采蛋可以为管理种群规模提供一个微调的调节盘,这样,与过度的体育狩猎(或以猎取野鸟为幌子的扑杀)相比,潜在的过度捕捞对种群数量的威胁会更小;图4)。从早期生命阶段(卵或幼鱼)收获是IPLC常用的策略,以微调或减少习惯收获的影响(Gudmundsson 1979, Hunter等人2000,Lyver等人2008)。
实施习惯管理的注意事项和潜在好处
支持IPLC与环境接触、产生和维护生态知识以及实行习惯管理的努力,可以改善当地的保护和社区生计(Popp等人2019年,Freitas等人2020年)。然而,要实现习惯管理的潜在好处,需要解决几个相关的注意事项。在这里,我们重点关注与ngangui Tahu和其他地方的IPLC的习惯管理相关的三个注意事项。
首先,改善IPLC对非IPLC集团拟议或正在进行的环境管理信息的获取,可以让习惯管理者做出知情的决定,这是可持续发展的关键(Armitage 2005, Akins等,2019)。例如,在本研究中,缺乏fgnz允许的天鹅运动狩猎的可用信息,造成了不确定性新西兰tiaki希望可持续地收获卵子的人。区域野鸟管理(目前通过FGNZ)可以通过更密切地监测天鹅存活率的基线率和允许死亡率来缓解这种不确定性,例如,通过建立一个带状恢复计划(Williams 1979, Sandercock 2006),并与新西兰tiaki.我们开发的模型可以在未来潜在的共同管理安排中帮助沟通新西兰tiaki以及新西兰皇家政府的代表,参与者可以分享天鹅数量和不同生态压力的信息。IPLC和非IPLC群体之间的这种合作和知识共享可以提高对社会-生态系统和环境管理可持续性的集体理解(Folke等人,2005年,Armitage等人,2009年,Popp等人,2019年,Wehi等人,2019年)。重要的是,环境管理方面的有效合作需要政府支持适应性政策,这些政策承认IPLC对资源和决策的权利,监测工作(例如,带恢复项目),并需要时间在不同群体之间建立信任(Folke等人,2005年,Akins等人,2019年)。IPLC和科学家(IPLC和非IPLC)之间的合作也可以提供一个途径,部分资助新知识的产生,例如,使用人口模型(Jones等人,2015年,McKechnie等人,2020年)和支持习惯管理(Ens等人,2016年,Popp等人,2019年,Freitas等人,2020年)。
其次,遏制人为驱动的环境退化对支持习惯管理很重要(Tang和Gavin 2016, Lyver等人2019b)。例如,在过去的半个世纪里,Te Waihora的农业污染阻碍了天鹅的主要食物来源——大型植物的生长(Lineham 1983, Gerbeaux 1989),并明显降低了天鹅在收获后产卵的倾向(Williams 1979, Herse et al. 2021)。因此,尽管有狩猎天鹅带来的压力,但Te Waihora的污染限制了当地ngaoi Tahu人可持续采蛋的能力,因为天鹅对采蛋的适应能力下降,一些采蛋者担心食用含有污染物的鸡蛋(Bataille 2020年,Bataille等人2021年)。随着时间的推移,在新西兰tiaki以及与Te Waihora的传统收割机接触可能会导致当地对天鹅以及它们对卵收获和其他压力的反应的认识的丧失。由于习惯收获和资源共享对许多IPLC的福祉至关重要(Tyrrell 2008年,Caillon等人,2017年,Freitas等人,2020年),包括毛利人(kawallu 2000年,Bataille 2020年,Bataille等人,2021年),习惯管理参与的减少可能对文化健康和多样性产生级联效应(Lyver等人,2019b, Fernández-Llamazares等人,2021年,Yletyinen等人,2022年)。因此,为了减轻这些威胁,我们敦促政府采取基于地点的环境管理政策,承认环境和文化福祉之间的联系(Gavin等人2015,Caillon等人2017,Akins等人2019,Lyver等人2019a)。
最后,改善环境治理中的社会公平对于实现习惯管理的潜在效益至关重要(Brondizio和Tourneau, 2016, ruu等人,2017)。忽视IPLC权利和价值的限制性政策限制了参与以及知识的持续生成和更新(Stephenson等人,2014年,Parlee等人,2018年)。此类政策可能导致不适当的保护方法,偏向于主导文化,反对IPLC (Moller 1996, Tyrrell 2008, Eichler和Baumeister 2018, Parlee等人2018,Guibrunet等人2021)。例如,新西兰法律充分保护大多数本土鸟类不受任何形式的掠食,而不管它们的保护地位或对毛利人的价值如何,但却将其他被运动猎人珍视的物种列为猎鸟,包括一种保护地位为<年代pan class="ob-q">”全国脆弱<年代pan class="ob-q">”(pārera或灰色的鸭子,阿拉斯superciliosa;新西兰政府1953,Williams 2013b, Wehi和Lord 2017, Lyver等人2019a)。即使是黑天鹅,一种经常被狩猎爱好者和农民射杀的物种,新西兰tiaki未经保护部长或区域FGNZ管理者的特别许可,不得批准传统的鸡蛋收获,这与上文所述的《条约》不一致,并为ngaji Tahu人的参与创造了障碍(Bataille 2020, Bataille等人2021)。先不谈对习惯采蛋的虚伪限制(在20世纪,政府授权的野鸟管理人员销毁、收集和销售天鹅蛋长达数十年,同时禁止ngangui Tahu的习惯采蛋;Lamb 1964年,Williams 1979年,Pauling和Arnold 2008年),该研究的结果表明,考虑到Te Waihora种群对AHY天鹅死亡率的敏感性远高于对鸟巢孵化成功的变化的敏感性,对习惯采蛋的长期审查是没有根据的。因此,实施习惯管理需要面对针对IPLC的文化偏见和权力不对称,并采取公平的政策,使IPLC能够以文化上适当的方式参与和保护其土地和资源(Pinkerton等人2014年,ruu等人2017年,Wehi和Lord 2017年)。
总之,我们强调,在环境管理中融入不同的文化和视角可以提高社会生态系统中的知识生产、适应性和可持续性,特别是如果个人和群体分享权力、建立信任并找到共同的工作基础,例如环境管理中的共同利益(Folke等人2005,Armitage等人2009,Tengö等人2014,Bataille等人2021)。然而,个人和群体之间在观点、优先事项、能力、期望和权威方面的差异可能会引发冲突,从而散播分裂和不信任,限制管理(Moller 1996, 1998, Coombes和Hill 2005, Brondizio等人2021)。避免这种困境对合作和可持续性非常重要,因此需要采取措施解决和调解冲突(Coombes和Hill 2005年,Brondizio等人2021年),包括承认IPLC的权利和愿望(联合国大会2007年,McGregor等人2020年,Reyes-García等人2022年),以及建立协作跨文化学习和能力建设倡议(Whyte 2013年,Tengö等人2014年,Tipa等人2016年)。扭转人类活动导致的生物多样性和生态系统健康下降显然并不容易,因此我们敦促各国政府和环境管理者寻求既有效又社会公平的适应性解决方案。
对本文的回应
欢迎对本文进行回复。如果被接受发表,您的回复将被超链接到文章。<一个href="//www.dpl-cld.com/responses.php?articleid=13410&mode=add">要提交回复,请遵循此链接.<一个href="//www.dpl-cld.com/responses.php?articleid=13410">要阅读已接受的回复,请按此链接.作者的贡献
所有作者都对研究的设计和稿件的撰写做出了贡献。M.R.H.负责数据收集、建模和手稿的撰写。N.J.S.和p.o.l获得了这项研究的主要资金。
致谢
我们感谢ngongi Tahu咨询委员会成员Donald Brown、iian Cranwell、Cyril Gilroy、John Henry、David Higgins(主席)、Jane Kitson、Craig Pauling、Makarini Rupene、Khyla Russell、Joe Taurima和已故Paul Wilson的专家建议和指导。我们感谢飞行员伊沃·约克尼和休·罗宾逊,船长兰尼·毕晓普,以及现场助理索菲·艾伦,阿尔巴·塞万提斯·洛雷托,Laís玛雅,米歇尔·马拉菲尼,奥斯卡·波拉德,马库斯·沙德博尔特和劳拉·杨的辛勤工作。克里斯·琼斯(Chris Jones)参与了这项研究的初步设计和开发。我们感谢Murray Williams提供的见解和信息,以及Lucas Pereira Martins和Brett Sandercock提供的帮助分析的R脚本。该项目由商业、创新和就业部的新西兰生物遗产国家科学挑战(项目3.2)和战略科学投资基金资助。
数据可用性
支持本研究结果的数据和R代码由Te rongnanga o ngongi Tahu和Manaaki Whenua Landcare Research联合持有。数据储存于MWLR (<一个href="https://doi.org/10.34721/HPRD-HY85" target="_blank" rel="noopener">https://doi.org/10.34721/HPRD-HY85).
文献引用
亚当斯,1971。埃尔斯米尔湖的黑天鹅。野生生物3:23-25审查。
阿金斯,p.o 'B。莱弗、H. F.阿尔øe和H.莫勒。2019。普遍预防原则:环境、社会文化和经济复原力的新支柱和新途径。可持续性11(8):2357。<一个href="https://doi.org/10.3390/su11082357" target="_blank" rel="noopener">https://doi.org/10.3390/su11082357
阿米蒂奇,d . 2005。适应能力和社区自然资源管理。环境管理35(6):703 - 715。<一个href="https://doi.org/10.1007/s00267-004-0076-z" target="_blank" rel="noopener">https://doi.org/10.1007/s00267-004-0076-z
阿米蒂奇,D. R.普卢默,F.伯克斯,R. I.亚瑟,A. T.查尔斯,I. J.戴维森-亨特,A. P.迪达克,N. C.杜布尔戴,D. S.约翰逊,M.马什克,P.麦康尼,E. W.平克顿,和E. K.渥伦伯格。2009。社会生态复杂性的适应性协同管理。生态环境科学进展7(2):95-102。<一个href="https://doi.org/10.1890/070089" target="_blank" rel="noopener">https://doi.org/10.1890/070089
Artelle, K. A, M. Zurba, J. Bhattacharyya, D. E. Chan, K. Brown, J. Housty和F. Moola. 2019。支持恢复土著主导的治理:公正和有效的保护新机制。生物保护240:108284。<一个href="https://doi.org/10.1016/j.biocon.2019.108284" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2019.108284
Ban, n.c., L. Eckert, M. McGreer和A. Frid. 2017。土著知识作为现代渔业管理的数据:以太平洋加拿大的邓杰内斯蟹为例。生态系统健康与可持续发展3(8):1379887。<一个href="https://doi.org/10.1080/20964129.2017.1379887" target="_blank" rel="noopener">https://doi.org/10.1080/20964129.2017.1379887
巴克尔,r.j., i.m.布坎南,1993。估计未成熟的新西兰黑天鹅的成活率。野生动物管理学报57(3):549-555。<一个href="https://doi.org/10.2307/3809281" target="_blank" rel="noopener">https://doi.org/10.2307/3809281
巴特,J. S.欧内斯特和P. J.培根。1991。天鹅的比较人口学研究进展。西尔斯和培根的15-21页,编辑。第三届国际天鹅研讨会论文集。野禽副刊第1号。国际水禽和湿地研究,斯利姆布里奇,英国。
巴塔耶,2020年。跨文化利益集团的价值观和期望状态为新西兰奥特里亚湿地的生物文化管理提供了信息。论文。坎特伯雷大学,基督城,新西兰。
巴塔耶,c.y, S. K.马利宁,J. Yletyinen, N. Scott, P. O 'B。Lyver》2021。关系价值为跨文化湿地管理提供了共同基础,揭示了多层次的约束。人与自然3(4):941-960。<一个href="https://doi.org/10.1002/pan3.10244" target="_blank" rel="noopener">https://doi.org/10.1002/pan3.10244
伯克,f . 2009。了解和研究环境变化的土著方式。新西兰皇家学会学报39(4):151-156。<一个href="https://doi.org/10.1080/03014220909510568" target="_blank" rel="noopener">https://doi.org/10.1080/03014220909510568
布拉德肖,c.j.a, B. W.布鲁克,2007。北澳大利亚一种濒危但外来的超级食草动物可持续收获的生态经济模型。自然资源建模20(1):129-156。<一个href="https://doi.org/10.1111/j.1939-7445.2007.tb00203.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1939-7445.2007.tb00203.x
Brockington, D.和J. Igoe. 2006。为保护环境而驱逐:全球概述。保护与社会4(3):424-470。
布罗迪齐奥,E. S., K.安德森,F.德卡斯特罗,C.普天玛,C.索尔克,M. Tengö, M.伦敦,D. C.图恩,T. S.冈萨雷斯,A. Molina-Garzón, G.罗素·洛佩斯,和S. M.西亚尼,2021年。在巴西亚马逊地区推行以地点为基础的可持续发展倡议。环境可持续性的观点49:66-78。<一个href="https://doi.org/10.1016/j.cosust.2021.03.007" target="_blank" rel="noopener">https://doi.org/10.1016/j.cosust.2021.03.007
布朗迪齐奥,e。S。和f。m。唐龙l .》2016。人人享有环境治理。科学》352(6291):1272 - 1273。<一个href="https://doi.org/10.1126/science.aaf5122" target="_blank" rel="noopener">https://doi.org/10.1126/science.aaf5122
布鲁克,B. W., D. M. J. S.鲍曼,C. J. A.布拉德肖,B. M.坎贝尔,P. J.怀特黑德。2006。在澳大利亚国家公园管理濒危的亚洲牛鼬:生态经济模型在决策中的作用和限制。环境管理38(3):463 - 469。<一个href="https://doi.org/10.1007/s00267-005-0157-7" target="_blank" rel="noopener">https://doi.org/10.1007/s00267-005-0157-7
汉堡,j . 1988。筑巢最少燕鸥的社会吸引力:数量、间距和成对关系的影响。秃鹰90(3):575 - 582。<一个href="https://doi.org/10.2307/1368345" target="_blank" rel="noopener">https://doi.org/10.2307/1368345
Caillon, S., G. Cullman, B. Verschuuren和E. J. Sterling. 2017。通过生物文化方法超越人与自然的二分法:在复原力指标中包括生态福祉。生态与社会22(4):27。<一个href="https://doi.org/10.5751/ES-09746-220427" target="_blank" rel="noopener">https://doi.org/10.5751/ES-09746-220427
卡斯韦尔,h . 2001。矩阵人口模型:构建、分析与解释。第二版。Sinauer Associates,桑德兰,马萨诸塞州,美国。
Chaloupka, M.和G. Balazs. 2007。利用贝叶斯状态空间模型评估夏威夷绿海龟种群的恢复和收获潜力。生态模型205(2):93 - 109。<一个href="https://doi.org/10.1016/j.ecolmodel.2007.02.010" target="_blank" rel="noopener">https://doi.org/10.1016/j.ecolmodel.2007.02.010
辛纳,J. E.和S.阿斯瓦尼。2007。将习惯管理融入海洋保护。生物保护140(3 - 4):201 - 216。<一个href="https://doi.org/10.1016/j.biocon.2007.08.008" target="_blank" rel="noopener">https://doi.org/10.1016/j.biocon.2007.08.008
库奇,g.g., M.吉列曼,G. S.布默尔,j . d。Lebreton和J. D. Nichols, 2014。收获对水禽种群的影响。野禽特刊第4期220-227。
库姆斯,B. L.和S.希尔,2005。<年代pan class="ob-q">”Na whenua, Na Tuhoe。Ko D.o.C. te合伙人<年代pan class="ob-q">”-特乌里韦拉国家公园管理前景。社会与自然资源18(2):135-152。<一个href="https://doi.org/10.1080/08941920590894516" target="_blank" rel="noopener">https://doi.org/10.1080/08941920590894516
卡明,G. S., D. H. M.卡明,C. L.雷德曼。2006。社会生态系统中的规模不匹配:原因、后果和解决办法。生态与社会11(1):14。<一个href="https://doi.org/10.5751/ES-01569-110114" target="_blank" rel="noopener">https://doi.org/10.5751/ES-01569-110114
Dawson, n.m., B. Coolsaet, E. J. Sterling, R. Loveridge, n.d. gros- camp, S. Wongbusarakum, K. K. Sangha, L. M. Scherl, H. Phuong Phan, N. Zafra-Calvo, W. G. Lavey, P. Byakagaba, C. J. Idrobo, A. Chenet, n.j. Bennett, S. Mansourian, F. J. Rosado-May。2021.土著人民和地方社区在有效和公平的保护方面的作用。生态与社会26(3):19。<一个href="https://doi.org/10.5751/ES-12625-260319" target="_blank" rel="noopener">https://doi.org/10.5751/ES-12625-260319
Díaz, S, J.塞特勒,E. S. Brondízio, H. T. Ngo, J. Agard, A. Arneth, P. Balvanera, K. A. Brauman, S. H. M. Butchart, K. M. Chan, L. A. Garibaldi, K. Ichii, J. Liu, S. M. Subramanian, G. F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R. R. Chowdhury, y - j。Shin, I. Visseren-Hamakers, K. J. Willis和C. N. Zayas, 2019。地球上普遍存在的人类导致的生命衰退表明需要进行革命性的变革。科学》366 (6471):eaax3100。<一个href="https://doi.org/10.1126/science.aax3100" target="_blank" rel="noopener">https://doi.org/10.1126/science.aax3100
埃奇勒,L.鲍迈斯特,2018。狩猎为正义:原住民对北美野生动物保护模式的批判。环境与社会9:75-90。<一个href="https://doi.org/10.3167/ares.2018.090106" target="_blank" rel="noopener">https://doi.org/10.3167/ares.2018.090106
Ens, E., M. L. Scott, Yugul Mangi Rangers, C. Moritz和R. Pirzl. 2016。将土著保护政策付诸实践可以带来生物多样性和文化效益。生物多样性与保护25(14):2889-2906。<一个href="https://doi.org/10.1007/s10531-016-1207-6" target="_blank" rel="noopener">https://doi.org/10.1007/s10531-016-1207-6
Ens, E., g.m. Towler, C. Daniels, Yugul Mangi Rangers, manqurk Rangers. 2012。回顾前进:在偏远的阿纳姆地进行协同生态监测。生态管理与修复13(1):26-35。<一个href="https://doi.org/10.1111/j.1442-8903.2011.00627.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1442-8903.2011.00627.x
埃扎德,T. H. G, J. M.布洛克,H. J.达格利什,A.米伦,F. Pelletier, A. Ozgul, D. N.昆斯。2010。变化世界的矩阵模型:瞬态动态在人口管理中的重要性。应用生态学报47(3):515-523。<一个href="https://doi.org/10.1111/j.1365-2664.2010.01801.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1365-2664.2010.01801.x
Fernandez-Llamazares,。,D. Lepofsky, K. Lertzman, C. G. Armstrong, E. S. Brondizio, M. C. Gavin, P. O’B. Lyver, G. P. Nicholas, P. Pascua, N. J. Reo, V. Reyes-García, N. J. Turner, J. Yletyinen, E. N. Anderson, W. Balée, J. Cariño, D. M. David-Chavez, C. P. Dunn, S. C. Garnett, S. Greening (La'goot), S. Jackson (Niniwum Selapem), H. Kuhnlein, Z. Molnár, G. Odonne, G.-B. Retter, W. J. Ripple, L. Sáfián, A. S. Bahraman, M. Torrents-Ticó, and M. B. Vaughan. 2021. Scientists’ warning to humanity on threats to Indigenous and local knowledge systems. Journal of Ethnobiology 41(2):144-169.<一个href="https://doi.org/10.2993/0278-0771-41.2.144" target="_blank" rel="noopener">https://doi.org/10.2993/0278-0771-41.2.144
C.福尔克,T.哈恩,P.奥尔森和J.诺伯格,2005。社会生态系统的适应性治理。环境与资源年度评论30:441-473。<一个href="https://doi.org/10.1146/annurev.energy.30.050504.144511" target="_blank" rel="noopener">https://doi.org/10.1146/annurev.energy.30.050504.144511
福特,D. E., F. D.休伊和K.泰勒,2017。Te Waihora/埃尔斯米尔湖:2017年的湖的状态。报告3号。Waihora Ellesmere信托技术公司,新西兰基督城。
Freitas, c.t, P. F. Macedo Lopes, J. V. Campos‐Silva, M. M. Noble, R. Dyball和C. A. Peres. 2020。共同管理具有重要文化意义的物种:促进生物多样性保护和人类福祉的工具。人与自然2(1):61-81。<一个href="https://doi.org/10.1002/pan3.10064" target="_blank" rel="noopener">https://doi.org/10.1002/pan3.10064
加吉尔,M., F. Berkes和C. Folke, 1993。保护生物多样性的土著知识。中22(3):151 - 156。
Gavin, m.c., J. McCarter, A. Mead, F. Berkes, J. R. Stepp, D. Peterson, R. Tang. 2015。定义生物文化保护方法。生态与进化动态30(3):140-145。<一个href="https://doi.org/10.1016/j.tree.2014.12.005" target="_blank" rel="noopener">https://doi.org/10.1016/j.tree.2014.12.005
杰博,1989。埃尔斯米尔湖水生植物的减少:新西兰一个浅湖的大型植物管理案例。论文。林肯学院,林肯,新西兰。
高德曼,2011。在自己的土地上的陌生人:坦桑尼亚北部的马赛人和野生动物保护。保护与社会9(1):64-79。
Gudmundsson, f . 1979。Mývatn水禽种群的过去状况和开发利用。Oikos 32(1 - 2): 232 - 249。<一个href="https://doi.org/10.2307/3544230" target="_blank" rel="noopener">https://doi.org/10.2307/3544230
格雷罗,a.m., R. R. J.麦卡利斯特,J.科克伦,K. A.威尔逊。2013。规模不匹配,保护计划,以及社会网络分析的价值。保护生物学27(1):35-44。<一个href="https://doi.org/10.1111/j.1523-1739.2012.01964.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1523-1739.2012.01964.x
吉布鲁内,l.p.。R. W. Gerritsen, J. A. Sierra‐Huelsz, A. C. Flores‐Díaz, E. García‐Frapolli, E. García‐serrano, U. Pascual和P. Balvanera. 2021。在参与之外:如何实现对当地社区保护价值体系的认可?以下是来自墨西哥的一些见解。人与自然3(3):528-541。<一个href="https://doi.org/10.1002/pan3.10203" target="_blank" rel="noopener">https://doi.org/10.1002/pan3.10203
哈利姆,A. A.奥斯曼,S. R.伊斯梅尔,J. A.贾万,N. N.易卜拉欣。2012。马来西亚沙巴的土著知识和生物多样性保护。国际社会科学与人文学报2(2):159-163。<一个href="https://doi.org/10.7763/IJSSH.2012.V2.88" target="_blank" rel="noopener">https://doi.org/10.7763/IJSSH.2012.V2.88
赫恩肖,E. J. S.和K. F. D.休伊。2010。Te Waihora/Lake Ellesmere提供的价值调查的容忍范围方法。土地环境与人研究报告第17号。林肯大学,林肯,新西兰。
先生,先生,p.o 'B。莱弗,N. J.斯科特,A. R.麦金托什,S. C.科茨,A. M.戈姆利,J. M.提利亚纳基斯,2020年。让土著人民和当地社区参与进来可以缓解社会生态系统的规模不匹配。生物科学70(8):699 - 707。<一个href="https://doi.org/10.1093/biosci/biaa066" target="_blank" rel="noopener">https://doi.org/10.1093/biosci/biaa066
赫斯、M. R.、J. M.提利亚纳基斯、N. J.斯科特、D.布朗、I.克兰韦尔、J.亨利、C.鲍林、A. R.麦金托什和P. O 'B。Lyver》2021。传统的采蛋制度对文化上重要的水禽种孵化成功的影响。人与自然3(2):499-512。<一个href="https://doi.org/10.1002/pan3.10196" target="_blank" rel="noopener">https://doi.org/10.1002/pan3.10196
汉恩,E. D.约翰逊,P.罗素,T.桑顿,2003。Huna Tlingit传统的环境知识,保护和管理<年代pan class="ob-q">”荒野<年代pan class="ob-q">”公园。当代人类学44(增刊):79 - 103。<一个href="https://doi.org/10.1086/377666" target="_blank" rel="noopener">https://doi.org/10.1086/377666
C.亨特,H.莫勒和J.基特森,2000。在新西兰收获的黑灰鹱(titi)雏鸟的选择性。新西兰动物学杂志27(4):395-414。<一个href="https://doi.org/10.1080/03014223.2000.9518249" target="_blank" rel="noopener">https://doi.org/10.1080/03014223.2000.9518249
生物多样性和生态系统服务政府间科学政策平台。2019 a。给决策者的生物多样性和生态系统服务全球评估报告摘要。S. Díaz, J.塞特莱,E. S. Brondízio, H. T. Ngo, M. Guèze, J. Agard, A. Arneth, P. Balvanera, K. A. Brauman, S. H. M. Butchart, K. M. Chan, L. A. Garibaldi, K. Ichii, J. Liu, S. M. Subramanian, G. F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R. Roy Chowdhury, Y. j Shin, I. J. Visseren-Hamakers, K. J. Willis, C. N. Zayas编辑。IPBES秘书处,德国波恩。<一个href="https://doi.org/10.5281/zenodo.3553579" target="_blank" rel="noopener">https://doi.org/10.5281/zenodo.3553579
生物多样性和生态系统服务政府间科学政策平台。2019 b。土著人民和当地社区。IPBES秘书处,德国波恩。<一个href="https://ipbes.net/glossary/indigenous-peoples-local-communities" target="_blank" rel="noopener">https://ipbes.net/glossary/indigenous-peoples-local-communities
琼斯,C. J., P. O 'B。莱弗、J.戴维斯、B.休斯、A.安德森和J.霍哈帕塔-奥克。2015.在暂停50年之后,重新恢复海鸟的常规捕捞。野生动物管理学报79(1):31-38。<一个href="https://doi.org/10.1002/jwmg.815" target="_blank" rel="noopener">https://doi.org/10.1002/jwmg.815
卡沃鲁,1989年。《怀唐伊条约》的翻译。川户主编,第321页。毛利人和巴基哈人对怀唐伊条约的看法。牛津大学出版社,奥克兰,新西兰。
Kawharu m . 2000。Kaitiakitanga:毛利人对资源管理的社会环境伦理的人类学视角。波利尼西亚学报109(4):349-370。
基南和P.达尔泽尔,2005。海龟作为一个旗舰物种:不同的观点在太平洋岛屿上制造了冲突。海事研究3(2):195 - 212。
昆斯,J. B.格兰德,B.津纳和R. F.洛克威尔,2005。瞬时种群动态:与生活史和初始种群状态的关系。生态模型185(2 - 4):283 - 297。<一个href="https://doi.org/10.1016/j.ecolmodel.2004.12.011" target="_blank" rel="noopener">https://doi.org/10.1016/j.ecolmodel.2004.12.011
昆斯,D. N., G. Gunnarsson, J. A. Schmutz和J. J. Rotella. 2014。水鸟种群动态的驱动因素:从蓝水鸭到天鹅。野禽特刊第4期169-191。
兰姆,1964年。鸟,兽和鱼:北坎特伯雷适应学会的前一百年。北坎特伯雷适应学会,基督城,新西兰。
莱斯利,p。h。1945。关于矩阵在某些人口数学中的应用。生物统计学33(3):183 - 212。<一个href="https://doi.org/10.1093/biomet/33.3.183" target="_blank" rel="noopener">https://doi.org/10.1093/biomet/33.3.183
Lineham, i . 1983。埃尔斯米尔湖富营养化:浮游植物的研究。论文。坎特伯雷大学,基督城,新西兰。
Lyver, p . O ' b。,J. Davis, L. Ngamane, L. Anderson, and P. Clarkin. 2008. Hauraki Maori Matauranga for the conservation and harvest of Titi, Pterodroma macroptera gouldi. Papers and Proceedings of the Royal Society of Tasmania 142(1):149-159.<一个href="https://doi.org/10.26749/rstpp.142.1.149" target="_blank" rel="noopener">https://doi.org/10.26749/rstpp.142.1.149
Lyver, p . O ' b。,J. Ruru, N. J. Scott, J. M. Tylianakis, J. Arnold, S. K. Malinen, C. Y. Bataille, M. R. Herse, C. J. Jones, A. M. Gormley, D. A. Peltzer, Y. Taura, P. Timoti, C. Stone, M. Wilcox, and H. Moller. 2019a. Building biocultural approaches into Aotearoa - New Zealand’s conservation future. Journal of the Royal Society of New Zealand 49(3):394-411.<一个href="https://doi.org/10.1080/03036758.2018.1539405" target="_blank" rel="noopener">https://doi.org/10.1080/03036758.2018.1539405
Lyver, p . O ' b。,P. Timoti, T. Davis, and J. M. Tylianakis. 2019b. Biocultural hysteresis inhibits adaptation to environmental change. Trends in Ecology & Evolution 34(9):771-780.<一个href="https://doi.org/10.1016/j.tree.2019.04.002" target="_blank" rel="noopener">https://doi.org/10.1016/j.tree.2019.04.002
Lyver, p . O ' b。,P. Timoti, S. J. Richardson, and A. M. Gormley. 2021. Alignment of ordinal and quantitative species abundance and size indices for the detection of shifting baseline syndrome. Ecological Applications 31(4):e02301.<一个href="https://doi.org/10.1002/eap.2301" target="_blank" rel="noopener">https://doi.org/10.1002/eap.2301
Lyver, p . O ' b。,而且J. M. Tylianakis. 2017. Indigenous peoples: conservation paradox. Science 357(6347):142-143.<一个href="https://doi.org/10.1126/science.aao0780" target="_blank" rel="noopener">https://doi.org/10.1126/science.aao0780
McGregor, D., S. Whitaker和M. Sritharan. 2020。土著环境正义和可持续性。环境可持续性的当前观点43:35-40。<一个href="https://doi.org/10.1016/j.cosust.2020.01.007" target="_blank" rel="noopener">https://doi.org/10.1016/j.cosust.2020.01.007
McKechnie, S, D. Fletcher, J. Newman, C. Bragg, P. W. Dillingham, R. Clucas, D. Scott, S. Uhlmann, P. O 'B。莱弗、A. M.戈姆利、拉基浦拉岛管理机构和H.莫勒,2020年出版。分离气候、副渔获物、捕食和收获对捕鲸的影响Ardenna盘菌)新西兰的人口动态:基于模型的评估。《公共科学图书馆•综合》15 (12):e0243794。<一个href="https://doi.org/10.1371/journal.pone.0243794" target="_blank" rel="noopener">https://doi.org/10.1371/journal.pone.0243794
说,t . 1988。北坎特伯雷的水禽管理。论文。坎特伯雷大学,基督城,新西兰。
穆勒,h . 1996。习惯使用土著野生动物——以双文化的方式保护新西兰的生物多样性。B.麦克费根和P.辛普森的89-125页,编辑。生物多样性:生物多样性系列研讨会的论文。新西兰惠灵顿自然保护部科学研究处。
穆勒,h . 1998。从互不信任到共同管理。新西兰渔业与狩猎通讯6:17-24。
莫勒,H. F.伯克斯,P. O 'B。Lyver和M. Kislalioglu, 2004。结合科学和传统生态知识:监测种群共同管理。生态与社会9(3):2。<一个href="https://doi.org/10.5751/ES-00675-090302" target="_blank" rel="noopener">https://doi.org/10.5751/ES-00675-090302
蒙太诺,W. F. D.范东恩,M. A.韦斯顿,R. A.穆德,R. W.罗宾逊,M.考林,p . j。Guay。2018。人类促成的黑天鹅在澳大利亚和新西兰殖民历史的基因评估。进化应用11(3):364 - 375。<一个href="https://doi.org/10.1111/eva.12535" target="_blank" rel="noopener">https://doi.org/10.1111/eva.12535
蒙太诺,W. F. D.范东恩,M. A.韦斯顿,R. A.穆德,R. W.罗宾逊,M.考林,p . j。Guay 2018 b。对罗伦斯等人(2017)的回应:本土还是非本土?灭绝和现存的新西兰黑天鹅的DNA。进化应用11(3):378 - 379。<一个href="https://doi.org/10.1111/eva.12587" target="_blank" rel="noopener">https://doi.org/10.1111/eva.12587
新西兰政府,1953年。野生动物法案1953。1953年第31号公共法令。新西兰政府,惠灵顿,新西兰。
新西兰政府,1987年。保护法案1987。1987年第65号公共法令。新西兰政府,惠灵顿,新西兰。<一个href="https://www.legislation.govt.nz/act/public/1987/0065/latest/DLM103610.html" target="_blank" rel="noopener">https://www.legislation.govt.nz/act/public/1987/0065/latest/DLM103610.html
新西兰政府,1998年。《1998年ngongi Tahu索赔解决法案》。公共法令1998年第97号。新西兰政府,惠灵顿,新西兰。<一个href="https://www.legislation.govt.nz/act/public/1998/0097/latest/whole.html" target="_blank" rel="noopener">https://www.legislation.govt.nz/act/public/1998/0097/latest/whole.html
橙色,c . 2011。怀唐伊条约。第二版。布里奇特·威廉姆斯书店,惠灵顿,新西兰。<一个href="https://doi.org/10.7810/9781877242489" target="_blank" rel="noopener">https://doi.org/10.7810/9781877242489
Parlee, B. L. J. Sandlos和D. C. Natcher. 2018。破坏生存:北美草原上的荒地驯鹿<年代pan class="ob-q">”开放获取的悲剧。<年代pan class="ob-q">”科学进步4 (2):e1701611。<一个href="https://doi.org/10.1126/sciadv.1701611" target="_blank" rel="noopener">https://doi.org/10.1126/sciadv.1701611
C.鲍林和J.阿诺德,2008。文化健康湖。K.休伊和K.泰勒的77-83页,编辑。Te Waihora/埃尔斯米尔湖:湖的现状和未来的管理。林肯大学,林肯,新西兰。
皮里,M. Z.贝克尔和S. R.贝辛格,2006。结合人口统计和基于计数的方法来识别受威胁海鸟的源汇动态。生态学应用16(4):1516 - 1528。<一个href="https://doi.org/10.1890/1051-0761(2006)016%5B1516:CDACAT%5D2.0.CO;2" target="_blank" rel="noopener">https://doi.org/10.1890/1051 - 0761 (2006) 016 (1516: CDACAT) 2.0.CO; 2
平克顿,E.安吉尔,N.拉德尔,P.威廉姆斯,M.尼科尔森,J.索尔克尔森和H.克利夫顿,2014。重建不列颠哥伦比亚省沿海渔业管理机构的地方和区域战略:管理的哪些组成部分是最关键的?生态与社会19(2):72。<一个href="https://doi.org/10.5751/ES-06489-190272" target="_blank" rel="noopener">https://doi.org/10.5751/ES-06489-190272
波普,J. N.普里亚德卡和C.科兹米克。2019。驼鹿共同管理的兴起和原住民知识的整合。野生动物的人类维度24(2):159-167。<一个href="https://doi.org/10.1080/10871209.2019.1545953" target="_blank" rel="noopener">https://doi.org/10.1080/10871209.2019.1545953
H. Pöysä, J.埃尔姆伯格,P.努米,K. Sjöberg。2004.可持续采伐的生态基础:补偿死亡率的普遍范式仍然有效吗?Oikos 104(3): 612 - 615。<一个href="https://doi.org/10.1111/j.0030-1299.2004.12788.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.0030-1299.2004.12788.x
R核心团队,2021年。R:统计计算的语言环境。R统计计算基金会,维也纳,奥地利。
罗伦斯,n.j., A.卡达马基,L. J.伊斯顿,A. J. D.丁尼森,R. P.斯科菲尔德,J. M.沃特斯。2017。古老的DNA和形态测量分析揭示了新西兰独特的黑天鹅的灭绝和更替。英国皇家学会学报B:生物科学284(1859):20170876。<一个href="https://doi.org/10.1098/rspb.2017.0876" target="_blank" rel="noopener">https://doi.org/10.1098/rspb.2017.0876
罗伦斯,n.j., A.卡达马基,L. J.伊斯顿,A. J. D.丁尼森,R. P.斯科菲尔德,J. M.沃特斯。2018。本地吗?古老的DNA否定了新西兰特有的黑天鹅的持久性:对蒙塔诺等人的答复。进化应用11(3):376 - 377。<一个href="https://doi.org/10.1111/eva.12577" target="_blank" rel="noopener">https://doi.org/10.1111/eva.12577
雷恩,A., G. Byrnes, L. Collier-Robinson, J. Hollows, A. McIntosh, M. Ramsden, M. Rupene, P. tamatii - elliffe, C. thomas和T. E. Steeves. 2020。以土著知识系统为中心,重新设想保护迁移。人与自然2(3):512-526。<一个href="https://doi.org/10.1002/pan3.10126" target="_blank" rel="noopener">https://doi.org/10.1002/pan3.10126
里德,J. M.和A. P.多布森。1993。行为约束与保护生物学:同质吸引与招募。生态与进化动态8(7):253-256。<一个href="https://doi.org/10.1016/0169-5347(93)90201-Y" target="_blank" rel="noopener">https://doi.org/10.1016/0169 - 5347 (93) 90201 - y
Reyes-Garcia V。,。Fernández-Llamazares, Y. Aumeeruddy-Thomas, P. Benyei, R. W. Bussmann, S. K. Diamond, D. García-del-Amo, S. Guadilla-Sáez, N. Hanazaki, N. Kosoy, M. Lavides, A. C. Luz, P. McElwee, V. J. Meretsky, T. Newberry, Z. Molnár, I. Ruiz-Mallén, M. Salpeteur, F. S. Wyndham, F. Zorondo-Rodriguez, E. S. Brondizio. 2022。确认土著人民和当地社区在2020年后生物多样性议程中的权利和作用。中51:84 - 92。<一个href="https://doi.org/10.1007/s13280-021-01561-7" target="_blank" rel="noopener">https://doi.org/10.1007/s13280-021-01561-7
鲁鲁,J., P. O 'B。莱弗,N. J.斯科特和D.埃德蒙兹,2017。扭转新西兰生物多样性下降的趋势:在改革后的保护法律中赋予马利人权利。政策季度13(2):65 - 71。<一个href="https://doi.org/10.26686/pq.v13i2.4657" target="_blank" rel="noopener">https://doi.org/10.26686/pq.v13i2.4657
Sandercock, B. K. 2006。从现场接触数据中估计人口统计参数:综述。野生动物管理学报70(6):1504-1520。<一个href="https://doi.org/10.2193/0022-541X(2006)70%5B1504:EODPFL%5D2.0.CO;2" target="_blank" rel="noopener">https://doi.org/10.2193/0022 - 541 x (2006) 70 (1504: EODPFL) 2.0.CO; 2
Sandercock, b.k, E. B. Nilsen, H. Brøseth, H. C. Pedersen. 2011。狩猎死亡率是自然死亡率的补充还是补偿?试验收获对柳松鸡存活和死因特异性死亡率的影响。动物生态学报80(1):244-258。<一个href="https://doi.org/10.1111/j.1365-2656.2010.01769.x" target="_blank" rel="noopener">https://doi.org/10.1111/j.1365-2656.2010.01769.x
谢尔,M. Boissière, G.博多安。2015。看不见的哨兵:保护盲区的局部监测和控制。生态与社会20(2):39。<一个href="https://doi.org/10.5751/ES-07625-200239" target="_blank" rel="noopener">https://doi.org/10.5751/ES-07625-200239
斯努克,J., A. Cunsolo, D. Borish, C. Furgal, J. D. Ford, I. Shiwak, C. T. R. Flowers和S. L. Harper, 2020年。<年代pan class="ob-q">”我们成为罪犯就是为了吃土地<年代pan class="ob-q">”:殖民地对野生动物的管理以及对因纽特人福祉的影响。可持续性12(19):8177。<一个href="https://doi.org/10.3390/su12198177" target="_blank" rel="noopener">https://doi.org/10.3390/su12198177
Sobrevila, c . 2008。土著人民在生物多样性保护中的作用:自然但经常被遗忘的伙伴。世界银行集团第44300号报告。世界银行集团,美国华盛顿特区。
所罗门,m . 2014。国家公园被封锁:来自Kaiwhakahaere taho Mark Solomon的行动号召。Te Karaka 63:14-17。<一个href="https://ngaitahu.iwi.nz/our_stories/locked-national-parks/" target="_blank" rel="noopener">https://ngaitahu.iwi.nz/our_stories/locked-national-parks/
斯蒂芬森,J., F. Berkes, N. J. Turner, J. Dick. 2014。海洋生态系统的生物文化保护:来自新西兰和加拿大的例子。印度传统知识杂志13(2):257-265。
斯塔本,C., B. Milligan和P. Nantel, 2020。矩阵种群模型的构建和分析,版本2.7,<一个href="https://cran.r-project.org/web/packages/popbio/index.html" target="_blank" rel="noopener">https://cran.r-project.org/web/packages/popbio/index.html
Tang R.和M. C. Gavin. 2016。对传统生态知识的威胁分类及保护对策。自然资源与社会14(1):57-70。
Te rongnanga o ngongi Tahu和自然保护部,2005年。Te Waihora联合管理计划:Mahere Tukutahi o Te WaihoraTe rongnanga o ngongi Tahu和新西兰克赖斯特彻奇自然保护部。<一个href="https://api.ecan.govt.nz/TrimPublicAPI/documents/download/2738434" target="_blank" rel="noopener">https://api.ecan.govt.nz/TrimPublicAPI/documents/download/2738434
Tengö, M., E. S. Brondizio, T. Elmqvist, P. Malmer,和M. Spierenburg. 2014。连接不同的知识系统以加强生态系统治理:多证据库方法。中43(5):579 - 591。<一个href="https://doi.org/10.1007/s13280-014-0501-3" target="_blank" rel="noopener">https://doi.org/10.1007/s13280-014-0501-3
蒂帕,G. R.哈姆斯沃斯,E.威廉姆斯和J. C.基特森,2016。将莫霍托兰加莫霍里人纳入淡水管理、规划和决策。P. G.杰利曼,T.戴维,C. P.皮尔森和J. S.哈丁的613-637页,编辑。新西兰淡水科学的进展。新西兰水文学会,惠灵顿,新西兰。
Tyrrell m . 2008。努纳维克因纽特人对加拿大北极地区白鲸管理的看法。人类组织67(3):322 - 334。<一个href="https://doi.org/10.17730/humo.67.3.47826252k0623352" target="_blank" rel="noopener">https://doi.org/10.17730/humo.67.3.47826252k0623352
联合国大会,2007年。《联合国土著人民权利宣言》一个RES / 61/295)。联合国大会,美国纽约。
Wehi, p.m., J. R. Beggs, T. G. McAllister。2019.Ka mua, Ka muri:将毛利人纳入新西兰生态。生态学杂志43(3):3379。
韦希,p.m., J. M.洛德。2017。在生态修复中纳入文化实践的重要性。保护生物学31(5):1109 - 1118。<一个href="https://doi.org/10.1111/cobi.12915" target="_blank" rel="noopener">https://doi.org/10.1111/cobi.12915
温鲍姆、K. Z、J. S.布拉舍尔斯、C. D.戈尔登和W. M.盖兹。2013。寻找可持续性:野生动物收成的评估落伍了吗?生态学通讯16(1):99 - 111。<一个href="https://doi.org/10.1111/ele.12008" target="_blank" rel="noopener">https://doi.org/10.1111/ele.12008
怀特,K. P. 2013。论传统生态知识作为合作概念的作用:一种哲学研究。生态过程2:7。<一个href="https://doi.org/10.1186/2192-1709-2-7" target="_blank" rel="noopener">https://doi.org/10.1186/2192-1709-2-7
威廉,S. I.和A. E.斯托里,2002。循环产卵前出勤对山羊草同步繁殖的影响。水鸟25(2):156 - 163。<一个href="https://doi.org/10.1675/1524-4695(2002)025%5B0156:IOCPAO%5D2.0.CO;2" target="_blank" rel="noopener">https://doi.org/10.1675/1524 - 4695 (2002) 025 (0156: IOCPAO) 2.0.CO; 2
威廉姆斯,m . 1977。发现黑天鹅的地点,天鹅座atvatusLatham,在新西兰的Whangape湖和Ellesmere湖上。澳大利亚野生动物研究:289-299。<一个href="https://doi.org/10.1071/WR9770289" target="_blank" rel="noopener">https://doi.org/10.1071/WR9770289
威廉姆斯,m . 1979。黑天鹅的现状和管理天鹅座atratus会莱瑟姆在埃尔斯米尔湖<年代pan class="ob-q">”太太<年代pan class="ob-q">”风暴,1968年4月。新西兰生态学杂志2:34-41。
威廉姆斯,m . 1981。新西兰的人口统计天鹅座atratus会人口。G. V. T.马修斯和M.斯玛特主编的147-160页。第二届国际天鹅研讨会论文集。国际水禽和湿地局,英国斯利姆布里奇。
威廉姆斯,m . 2013 a。黑天鹅|卡卡普那瑙。c·m·米斯凯利,编辑。新西兰鸟类在线。<一个href="https://nzbirdsonline.org.nz/species/black-swan" target="_blank" rel="noopener">https://nzbirdsonline.org.nz/species/black-swan
威廉姆斯,M. 2013b[更新2022]。灰鸭|帕雷拉。c·m·米斯凯利,编辑。新西兰鸟类在线。<一个href="https://nzbirdsonline.org.nz/species/grey-duck" target="_blank" rel="noopener">https://nzbirdsonline.org.nz/species/grey-duck
维特,R., K. R. M. Suiseeya, R. L. Gruby, S. Hitchner, E. M. Maclin, M. Bourque和J. P. Brosius. 2015。影响全球环境治理的时刻。环境政治24(6):894 - 912。<一个href="https://dx.doi.org/10.1080/09644016.2015.1060036" target="_blank" rel="noopener">https://dx.doi.org/10.1080/09644016.2015.1060036
伍德,K. A, L.曹,P.克劳森,C. R.伊利,L. Luigujõe, E. C.里斯,J.斯奈德,D. V.索洛维耶娃,和R.沃达尔奇克。2019.天鹅研究的当前趋势和未来方向:来自第六届国际天鹅研讨会的见解。野禽特刊5:1-34。
Yletyinen, J. M. Tylianakis, C. Stone, P. O 'B。Lyver》2022。环境变化和政策对土著文化的潜在连锁影响。中51:1110 - 1122。<一个href="https://doi.org/10.1007/s13280-021-01670-3" target="_blank" rel="noopener">https://doi.org/10.1007/s13280-021-01670-3
Zador, s.g., J. F. Piatt和A. E. Punt. 2006。殖民地海鸟的捕食和卵收获平衡:一个模拟模型。生态模型195(3 - 4):318 - 326。<一个href="https://doi.org/10.1016/j.ecolmodel.2005.11.002" target="_blank" rel="noopener">https://doi.org/10.1016/j.ecolmodel.2005.11.002

图1

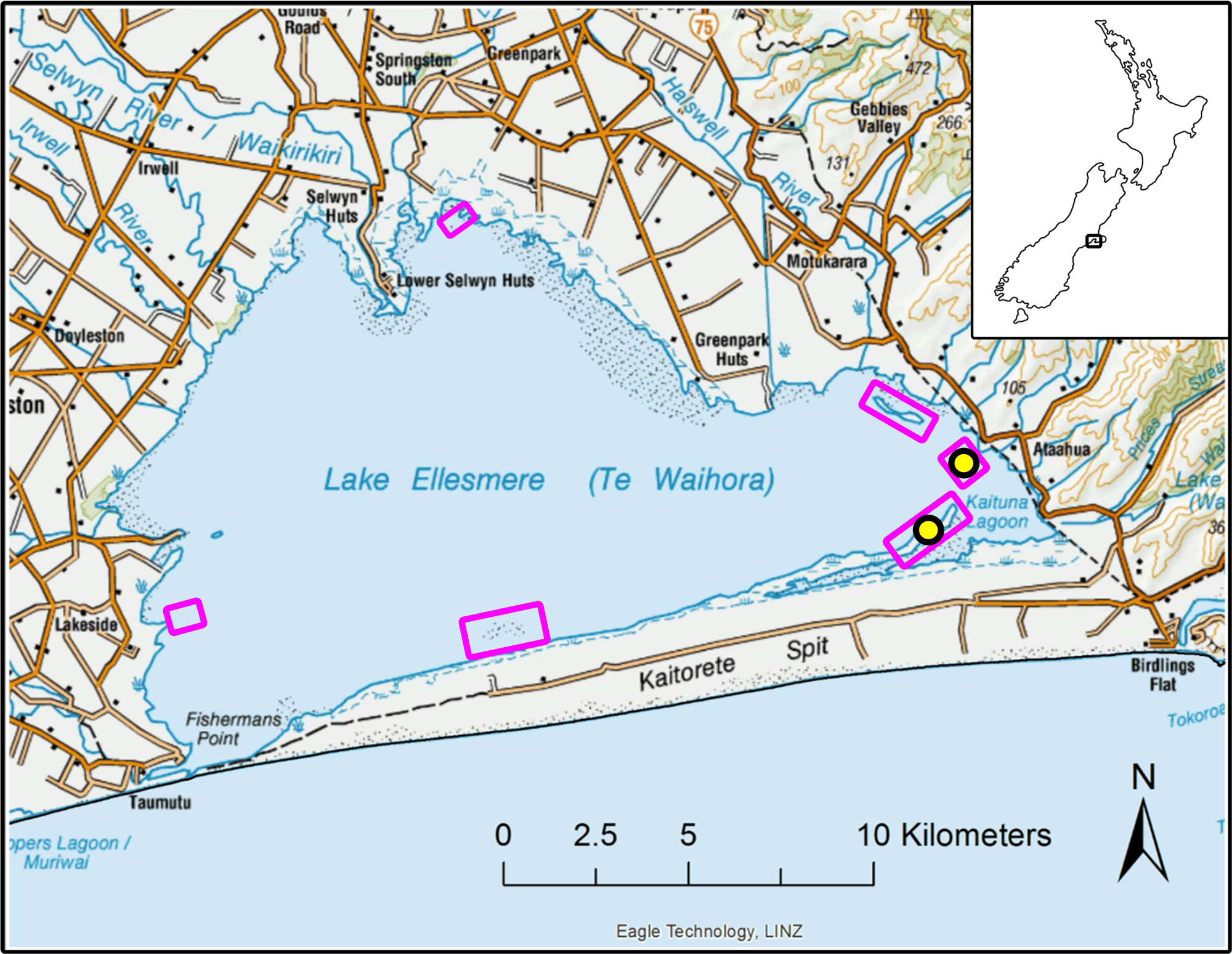
图1.我们在特怀霍拉/埃尔斯米尔湖的研究区地图,这里是黑天鹅数量最多的地方(天鹅座atratus会)在新西兰的南岛。紫色的方框勾勒出天鹅繁殖区,我们用正射影摄影法在那里普查巢穴。黑圈和黄圈表示Te Waihora最大的两个天鹅繁殖群的当前位置,我们在模型中使用的巢穴生育和卵子收获数据来自那里。该地图是在ArcGIS 10.5(环境系统研究所,Redlands, CA, USA;©鹰科技集团有限公司)。
图2
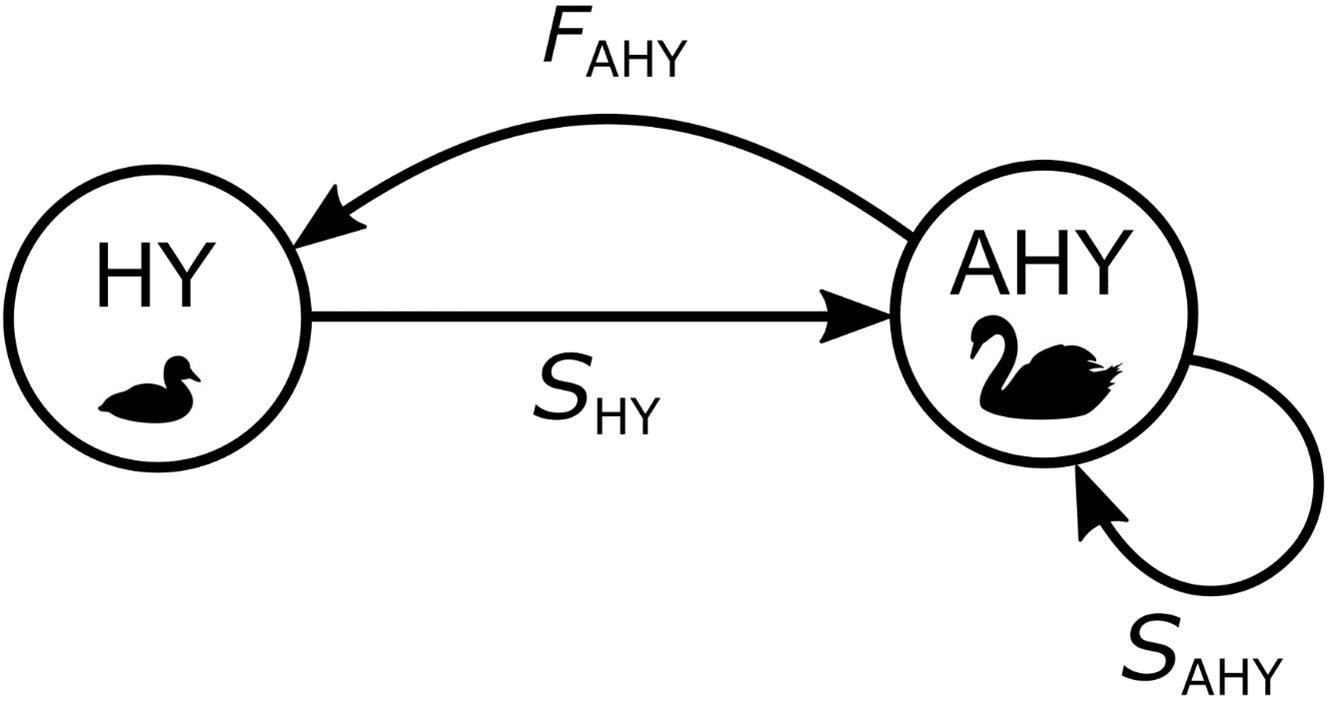
图2.描述黑天鹅的年龄分类矩阵模型的生命周期图(天鹅座atratus会)的人口动态在Te Waihora/Lake Ellesmere, Aotearoa/新西兰。圆圈表示孵化年龄(HY =孵化年,年龄< 1岁;AHY =孵化年后,年龄≥1岁),箭头表示年人均人口转换率(FAHY =年繁育率或每只AHY天鹅孵化的卵数;SHY =孵化后第一年存活的概率;SAHY = AHY天鹅的年生存概率)。
图3
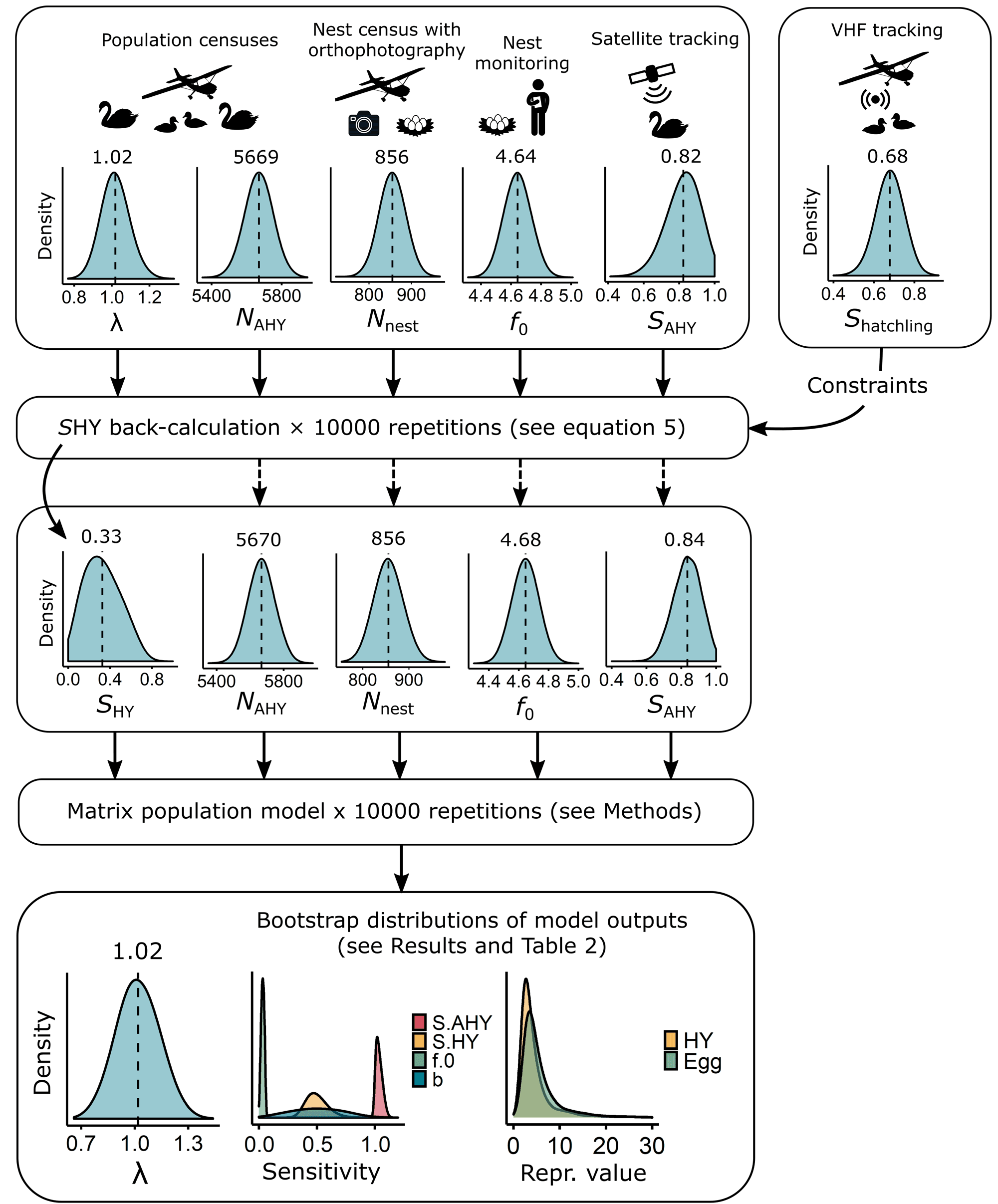
图3.流程图说明我们在开发矩阵模型和估计黑天鹅(天鹅座atratus会)的人口在Te Waihora/Lake Ellesmere, Aotearoa/新西兰。首先,我们使用经验数据定义了种群增长(λ)的初始自举抽样分布、每孵化年后天鹅年繁殖力(FAHY)的基本参数和AHY天鹅年生存概率(SAHY)。其次,我们使用初始分布中的随机抽样进行逆向计算,从而生成天鹅在其第一个(孵化)年的生存概率(SHY)的bootstrap分布。反向计算的SHY包含了天鹅孵化的生存信息(Shatchling),并调整了SAHY的分布和FAHY的基础参数。最后,我们从得到的自举分布中使用随机抽样来参数化矩阵模型,并为焦点总体的一组人口参数生成自举分布。
图4
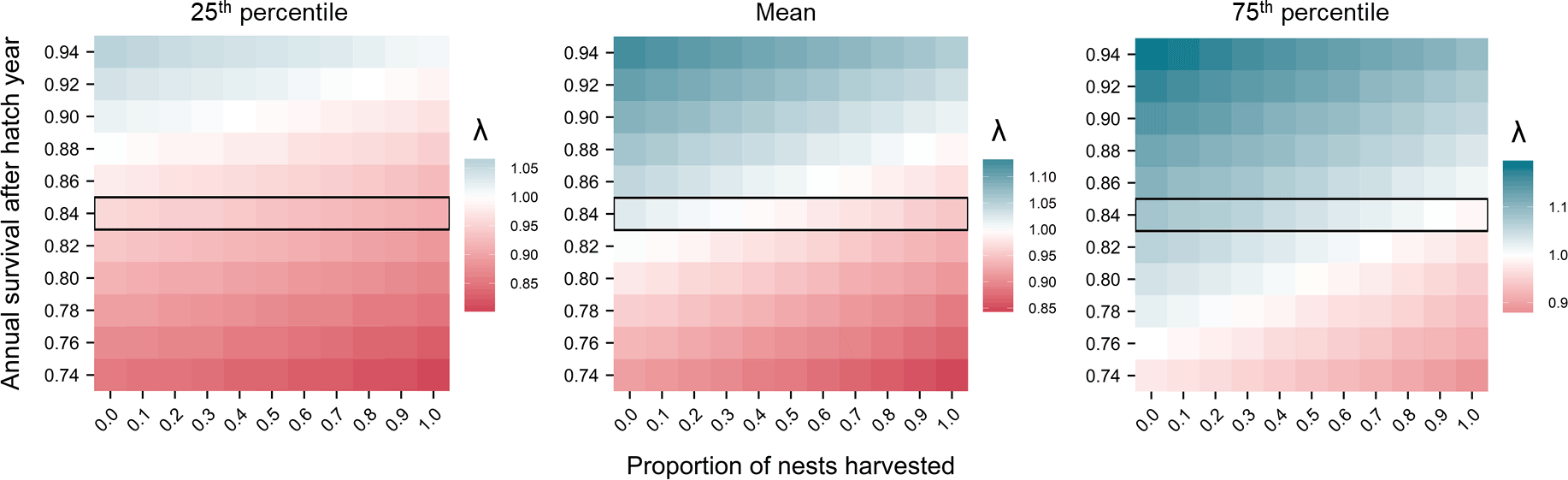
图4.热图显示了孵化年之后黑天鹅的年生存概率的变化(SAHY,天鹅座atratus会)和巢收获压力(h),影响Te Waihora/Lake Ellesmere, Aotearoa/New Zealand的种群增长率(λ)。暖色表示种群减少(λ < 1),冷色表示种群增长(λ > 1),中性白色表示种群稳定(λ = 1)。中间的图显示了SAHY和h的每个组合的λ自举分布的平均值,而左边和右边的图分别显示了每个组合的第25和第75百分位。
表1
表1.用于模拟黑天鹅(天鹅座atratus会)的人口动态在Te Waihora/Lake Ellesmere, Aotearoa/新西兰。人口统计率包括种群增长率(λ),每未收获的巢(f0)和每收获的巢(fa)的繁殖力(孵出的蛋数),总巢数(Nnest),孵化年后或AHY天鹅总数(NAHY),天鹅孵化头40天的生存概率(Shatchling),第一个(孵化)年的生存概率(SHY)和AHY天鹅的年度生存概率(SAHY)。用下标“a”标记的分布来反计算SHY,并根据与Shatchling相关的约束进行调整(见正文)。标下标“b”的分布被用来参数化矩阵模型,而标星号的分布被调整,但没有在矩阵模型中使用。
| 人口率 | Inititial引导分布 | 新的或调整的引导分配 |
| λ | 1.020 (0.890, 1.170)<年代up>一个 | 1.017 (0.898, 1.150)<年代up>* |
| f0 | 4.64 (3.55, 5.83)<年代up>一个 | 4.68 (3.60, 5.88)<年代up>b |
| f一个 | 2.67 (1.64, 3.83)<年代up>b | - |
| N巢 | 856(800、913)<年代up>一个 | 856(799、912)<年代up>b |
| NAHY | 5669(5521、5815)<年代up>一个 | 5670(5520、5815)<年代up>b |
| 年代人工孵化的 | 0.68 (0.54, 0.80)<年代up>一个 | 0.68 (0.54, 0.80)<年代up>* |
| 年代沪元 | - | 0.33 (0.06, 0.66)<年代up>b |
| 年代AHY | 0.82 (0.65, 1.0)<年代up>一个 | 0.84 (0.71, 0.94)<年代up>b |
表2
表2.与人口统计率相对应的不同参数的bootstrap分布的均值和相关的95%置信极限(括号中)(f0为生育力或每个未收获巢孵化的蛋数;B为年繁殖倾向;SHY是存活第一个或孵化年的概率;SAHY是确定黑天鹅(天鹅座atratus会)在Te Waihora/Lake Ellesmere /新西兰的人口增长(λ)。“敏感性”和“弹性”分别是某一人口比率中绝对变化和比例变化对λ的影响的量度。为了评估λ的不确定性来源,我们量化了每个人口比率的方差对λ的总方差的“贡献”。“繁殖价值”指的是单个AHY天鹅在其一生中平均产生的新鲜蛋和孵出的蛋(即HY天鹅)的总数量的估计。所有的估计都是基于一个模型,假设没有鸡蛋收获压力(h = 0);模型的这种特殊情况导致f0, b和SHY的弹性是相同的。
| 人口率 | 灵敏度的λ | 弹性λ | 贡献 不确定性λ |
| f0 | 0.032 (0.008, 0.056) | 0.145 (0.041, 0.229) | 0.027 |
| b | 0.502 (0.120, 0.886) | 0.145 (0.041, 0.229) | 0.001 |
| 年代沪元 | 0.499 (0.361, 0.694) | 0.145 (0.041, 0.229) | 0.530 |
| 年代AHY | 1.037 (1.002, 1.097) | 0.855 (0.771, 0.959) | 0.442 |