






以下是引用这篇文章的既定格式:
巴赫,J. Minderman, N. Bunnefeld, A. C. Mill和A. B. Duthie, 2022。干预还是等待?不确定性环境下保护冲突适应性管理干预时机评估模型。生态与社会27(3):3。摘要
生物多样性管理者干预的时机对保护的成功至关重要,特别是在保护目标与人类生计之间存在冲突的情况下,即保护冲突。考虑到与复杂的社会生态系统相关的不确定性,以及延迟行动对生物多样性和生计的潜在不可逆转的后果,管理者倾向于通过预防措施尽快进行干预。然而,在情况允许的情况下避免干预可能是有益的,特别是通过节省关键的管理资源。我们引入了一种策略,让管理人员根据监测来决定是否需要干预,或者是否可以等待。本研究评估了这种等待策略与无条件干预策略的性能。我们建立了一个以个体为基础的保护冲突模型,目的是保护动物种群的管理者和目的是通过保护他们的作物不受野生动物伤害来实现产量最大化的农民之间的保护冲突。然后,我们模拟了一种预算受限的适应性管理,随着时间的推移应用每种策略,同时考虑到人口动态和管理者和农民决策的不确定性。我们的研究结果表明,当管理者的干预决策是基于对种群轨迹的预测时,等待策略的表现至少与无条件干预一样好,同时也允许管理者通过避免不必要的干预来节省资源。在管理人员面临的困难预算限制下,这种等待策略确保了无条件干预的高收益,同时通过用等待期间积累的收益补偿管理人员缺乏资源,显著改善了保护结果。这表明,在保护冲突中,等待策略是值得考虑的,因为它可以促进公平管理,更有效地利用管理资源,而管理资源往往是生物多样性保护的限制因素。的简历
介绍
随着人口的增长和生活水平的提高,用于人类活动的地球表面的面积越来越大,经常与需要保护的物种的范围重叠。当这种物种受到严格保护,但又对人类生计产生影响时,就会出现保护冲突,可能导致管理决策的利益冲突(Redpath et al. 2013, 2015)。偏离目标可能导致土地使用者通过忽视或颠覆政策而背离政策,并从事往往阻碍保护目标的非法活动(Bunnefeld等人2013年,Bainbridge 2017年,Glynatsi等人2018年,Rakotonarivo等人2020年)。当养护和保护干扰到农业等基本生计活动时,这些冲突尤其严重(Behr等,2017,Mason等,2017)。因此,保护政策必须符合土地使用者的利益,以确保遵守和最大化保护成功,同时最大限度地减少对粮食安全和(或)农民收入的影响。此外,由于保护冲突形成了具有多种生物、环境、地理和社会成分的复杂系统,这些相互关联的社会-生态系统(SES)对变化的响应难以预测(van Wilgen and Biggs 2011, Game et al. 2013, Mason et al. 2018)。为了避免可能危及生物多样性保护或人类生计的不可预见的扰动,管理还应接受围绕生态过程和人类行为的不确定性(Fryxell等人2010,Bunnefeld等人2011,Schlüter等人2012,Cusack等人2020)。
应对不确定性挑战和复杂系统的一种实用方法是自适应管理,这是一种通过从结果中学习来迭代改进管理的技术(Williams et al. 1996, Hicks et al. 2009, Keith et al. 2011)。它特别适合于保护冲突管理,因为定期监测和政策更新增强了对立利益之间权衡的能力(Redpath et al. 2013, Wam et al. 2016, Mason et al. 2018, Richardson et al. 2020)。因此,适应性管理使保护政策尽可能贴近系统的变化,然而,何时以及为何更新政策是更好地管理SES和保护冲突的关键(Pérez等人,2019)。因为管理不善的后果可能是有害的,有时甚至是不可逆转的(例如,作物损失和/或动物种群灭绝;Kaswamila et al. 2007),传统智慧可能认为,尽可能多地对更新的政策作出反应将使保护的成功最大化。但如果计划得当,等待最终可以带来更好的管理结果,因为它可以带来各种好处,包括通过监测或研究提高知识(Walters 1986, Gregory等人2006,Nicol等人2018)。例如,Sims和Finnoff(2013)模拟了入侵物种缓慢而可预测的传播过程,并表明,由于在等待期间获得的知识,在减少对重点生态系统的传播和损害方面,首次干预的延迟时间比检测到入侵后立即进行干预更有效。相比之下,当入侵迅速而不稳定时,延迟的干预会导致对物种发展的控制失去,最终导致一种状态,在这种状态下,任何干预都变得毫无意义。在适应性管理背景下,Iacona等人(2017)对国家公园的鸟类多样性保护计划进行了建模,并表明,在将保护资金逐步用于保护之前,等待和储蓄保护资金以产生利息,可以实现更多的受保护物种数量和更快的灭绝债务的恢复。由于用于管理的财政和人力资源往往是有限的(huhey et al. 2003, McDonald-Madden et al. 2008),在保护资金分配限制允许的情况下,等到收益大于风险时可以避免不必要的支出(Ruiz-Miranda et al. 2020, Wu et al. 2020)。 This trade-off between instances of intervention and waiting in an adaptive management process has not yet, to our knowledge, been explored in the context of conservation conflicts. We hypothesize that by refraining from intervening when conflicting stakeholder interests are already aligned, managers could save resources and use them to enhance impact when intervention will be most needed to deliver conservation and/or land users’ objectives. We predict that it is likely to be especially relevant in situations in which a manager’s lack of resources could be compensated for by benefits accumulated over a period of waiting.
为了研究干预时机对管理质量的影响,同时考虑与保护冲突相关的不同不确定性来源,我们使用了广义管理策略评价框架(GMSE;Duthie等人,2018年)。广义管理战略评价框架建立在管理战略评价(MSE)框架的基础上,该框架探索了替代管理方案的可能结果,以评估它们对管理者目标的充分性(Smith et al. 1999)。管理战略评价首先在渔业领域发展起来,后来用于陆生物种,它利用种群动态、监测、管理决策和捕捞活动的子模型对自然资源适应管理的过程进行长期分解,这些子模型相互通知和影响。在评估场景时,这种结构有助于隔离与每个过程相关的不确定性的不同组成部分(Bunnefeld et al. 2011)。广义管理战略评估框架对所有四个子模型使用基于个体的方法,内在地模拟不确定性(Grimm 1999, DeAngelis和Grimm 2014),并包括一个决策子模型,模拟管理者和农民代理的目标导向行为,具有次优选择的可能性(遗传算法;Hamblin 2012, Duthie等人2018)。此外,通过产生代理之间的差异,基于个体的模型(ibm)可以模拟另一个潜在的冲突来源:成本和利益在利益相关者之间的不公平分配。Rakotonarivo等人(2020年,2021年)表明,农民对保护措施的更高公平感增加了选择支持保护的选项的倾向。因此,在保护冲突管理中,对用户间的权益进行建模和监测是非常重要的。 Knowing this, we further develop and apply GMSE to evaluate the efficiency of alternative management timing strategies against unconditional intervention and determine whether and how a profitable timing trade-off can be found for conservation conflict management under uncertainty.
我们建立了一个预算受限的保护冲突适应性管理模型,在该模型中,受保护关注的野生动物种群对农业活动有负面影响,农民可以通过扑杀来应对,以减少产量损失。我们为管理者提出了两种新的时间策略,以确定当通过等待节省的资源产生长期效益时,是否需要进行干预。通过GMSE模拟,我们评估了每种时间策略如何影响生物多样性保护与农业生产目标之间冲突的管理质量。因此,我们确定了在哪些情况下,我们的替代策略会比在每个机会都进行干预产生更好的管理效果。
方法
模型概述
模型中
为了模拟一段时间内的保护冲突管理,我们开发了一个基于个体的野生动物种群模型(称为”人口”)、农民和管理者都在一个农业景观上相互作用。景观被分割成离散的细胞,每个细胞产生农业产量,可以容纳任意数量的动物。每个农民都有一个连续的细胞块,形成他们的”土地,”细胞的生产力总和决定了农民的产量。动物为了生存和繁殖,从景观细胞中消耗农业资源,从而降低了农民的产量。农民可以扑杀自己土地上的动物,以减少产量损失。管理器试图通过将种群维持在预定义的目标大小(TN),就像以前在处理埃塞俄比亚山区尼亚拉羚羊保护和trophy hunting之间的冲突(Bunnefeld等人,2013年),或苏格兰或瑞典农业和候鸟保护之间的冲突(Bainbridge, 2017年,Mason等人,2017年,Nilsson等人,2021年)中所做的那样。选择这个目标是为了使其足够高以防止物种灭绝,但又足够低以确保农民获得满意的产量。管理者的方法是实施适当的政策,激励或抑制扑杀,使种群规模更接近TN.因此,在自适应管理过程之后,管理员根据对种群大小的监视更新此策略(Nt)在每个时间步t.
经理决策
经理收到固定的、非累积的预算(B米),我们将其解释为反映经理在实施政策更改和实施扑杀限制时可用的总时间、精力或金钱。该政策的模型是农民在他们的土地上扑杀动物时必须支付的成本。经理可以拉拢B米提高成本以阻止农民扑杀有利于人口增长,也可以降低成本以促进扑杀有利于人口减少。为了建立执行限制政策所需的预算模型,扑杀成本每增加1,就需要经理投入10个预算单位(b.u)。相反,当政策允许高扑杀率时,管理者不需要激励农民移走动物,他们不需要花费预算来降低成本。根据管理者的目标,用GMSE的进化算法计算管理者对筛选成本的改变量,该目标被建模为最小化两者之间的距离Nt而且TN鉴于农民的扑杀率与之前的成本相对应。
时机策略
我们探讨了三种定时策略,它们决定了管理人员是干预并更新策略,还是等待并保持策略不变。控制策略(CTL)为本研究的零模型。它对应于每一个机会的无条件干预,被建模为管理者在每一个时间步骤更新策略。利用自适应干预策略(ATI),我们定义了一个允许范围(PT周围)TN以TN±PT.在这个范围内,经理会考虑Nt足够接近TN,因此,决定了当前政策的结果是一个可持续的扑杀率的人口。因此,当且仅当在外部监视人口时,经理将更新策略TN±PT.轨迹策略(TRJ)与ATI策略相同,除了当Nt是内在的TN±PT,管理员根据当前和先前的监测结果,以线性外推的形式对下一个时间步的种群规模进行预测。如果这个预测是错误的TN±PT,经理保持政策不变;否则,他们会更新它。在ATI和TRJ策略中,在没有更新策略的时间步长之后,经理会获得额外的比例(预算奖金)Bb)的初始预算,以模拟与等待相关的好处。这种奖励可以在几个连续的等待时间步骤中积累,但当管理人员再次使用预算提高限制级别时,这种奖励就会丢失(建模细节见附录1)。
农民行动计划
在每个时间步的开始,每个农民都收到一个固定的、非累积的预算(BF),他们用这笔钱在自己的土地上宰杀一定数量的动物,费用由管理者的政策规定。一个农民扑杀一只动物的最低成本是10磅。使用GMSE的进化算法,每个农民被扑杀的牲畜数量都是独立计算出来的,这意味着每个农民都可以根据自己的目标(产量最大化)独立决定如何行动。
基于广义管理战略评价框架的仿真
为了模拟不确定性条件下不同时间策略下的保护冲突自适应管理,我们使用了R包GMSE (Duthie et al. 2018)。有关建模、参数选择和模拟的详细信息,请参见附录1。
初始参数
我们将景观建模为由40块大小相同的矩形土地组成的网格,每一块土地都属于一个农民。我们模拟了一个种群,它在没有扑杀的情况下是稳定的,但在高扑杀率下面临灭绝的威胁。我们确定了种群动态模型参数,使其快速稳定地达到均衡规模。当动物数量达到平衡时,为农民提供了足够高的初始预算,以在他们的土地上扑杀预期数量的动物(BF= 1000 b.u),所以如果冲突得不到控制,种群就会灭绝。起初,经理的初始预算被设定为与农民的预算相等(B米=BF= 1000 b.u),经理的目标设定为均衡大小的一半(TN= 2000只动物)。初始种群大小设为N0= 1000只动物,这个数字已经足够低,足以使动物数量面临立即灭绝的威胁,并证明管理者最初的介入是合理的。我们为控制策略选择这些参数,以产生足够的管理,同时也留下改进的空间,并确定替代策略可以产生更好的结果的程度。
种群动态被
广义管理战略评价的人口动态模型以人口为特征N每一种动物都有自己的年龄和位置。在每一个时间步中,每个动物都从它当前的细胞移动到一个特定范围内的随机细胞。到达后,动物吃掉细胞剩余产量的0.5%。所有的动物都在一个时间步长内以随机的顺序移动12次。在所有的运动和喂养之后,动物每消耗5个资源单位就会产生一个后代,这些后代作为新个体添加到种群中。接下来,那些消耗了超过四分之四的资源单位且年龄≤5个时间步长的动物存活下来,其余的动物将从种群中删除。这种消费标准导致了密度依赖性的种内资源竞争,对生活事件的离散建模产生了个体间的变异性以及地理和人口的随机性,因此解释了围绕种群动态的几个不确定性来源。
监控被
我们假设管理器在监视期间不会出错,因此Nt表示每个时间步上的确切总体大小。这一假设避免了建模的随机监测误差,这些误差会挑战对管理动力学的全面理解。
决策被
在每个时间步中,管理者和农民的决策使用进化算法独立建模,当agent的目标相反时,允许出现冲突。这种方法计算实际但不一定是最优的决策,认识到大多数人不能想到每一个选择最优的可能性,但可以在他们能想到的选项中选择最佳的选项(Hamblin 2012, Duthie et al. 2018),对利益相关者的个人决策产生不确定性。
实验计划
系统参数的探索
为了从人口动态和农民产量的角度评估ATI和TRJ的管理质量,我们改变了允许度PT和预算奖金Bb并将结果与CTL的结果进行比较。PT从经理目标的0%开始TN(无条件更新在每个时间步,即CTL)到100%TN(只在种群灭绝或在没有扑杀的情况下接近自然平衡的极端情况下更新)增加10%。Bb从经理初始预算的0%开始B米(等待时间后没有奖励)到100%B米10%的增量。对于每一个独特的组合PT而且Bb,在相同的初始条件下,我们在20个时间步骤中运行了100个独立的管理模拟重复。
管理的结果
我们将最理想的结果定义为当管理防止种群灭绝(1),同时使其尽可能接近目标(2),并确保农民的最高产量(3)和最低的不平等(4)。对于特定的参数组合,灭绝风险(1)被评估为在所有重复中灭绝事件的频率,记为fext.我们用种群大小(Nt)和经理的目标(TN)加权TN在模拟结束时,对所有重复的平均值表示dT,在%TN.农民的总产量(3)计算为模拟结束时所有细胞的产量总和与在没有动物消耗的情况下景观能提供的最大产量(40000个产量单位)的比值,该比值为所有重复的平均值并表示Y结束在景观的最大生产力。。农民之间的不平等(4)是用模拟结束时最高和最低的农民产量与最高产量之间的差值进行加权,表示为所有重复的平均值Yineq,在%的产量最高。最后,我们计算了没有管理者干预的时间步骤占模拟时间长度的比例,并在所有重复中取其平均值,表示tw(1 -tw因此是策略更新的比例)。我们在每个平均值周围计算了95%的自引导置信区间(Manly 2007)。通过系统地面对保护和农业成果来评估利益相关者之间的公平,以发现成本和收益的不平衡再分配。
对经理预算敏感
我们假设预算奖金数额(Bb)对管理质量的影响在预算约束较高的情况下会更强。为了验证这一点,我们选择了50%的许可度,TRJ的结果与CTL没有区别,但与弱Bb的效果。我们减少了经理的初步预算(B米)从1000磅增加到500磅,增加100磅。为每一个B米,我们不同Bb从0到100%B米在100个重复中增加10%,并测量与前一节相同的结果代理,以调查的影响Bb金额按管理质量B米.我们还模拟了每种CTL的管理B米值,以检查在比较中等待策略的执行情况。
结果
干预策略的自适应时机
保护的结果
当采用自适应时机干预(ATI)策略时,增加允许度值会导致物种灭绝风险增加,最终种群规模下降到目标以下,而预算红利(Bb;图1和附录2,图A2.1)。与CTL策略相比,允许度和奖励量的组合均不会导致相同或更低的灭绝风险(fext= 0.15 with [0.08;0.22] 95%置信区间)。ATI策略的参数组合均没有CTL策略更接近或同样接近目标(dT= -24.90% (-33.78;-16.26]),这并不奇怪,因为对于大多数组合来说,灭绝几乎是肯定的(fext> 0.9PT> 20%)。
农业的结果
增加许可程度导致农民的最终产量增加,农民之间的产量不平等减少,而预算奖金金额没有影响(附录2,图。A2.2和A2.3)。农户最终产量为ATI各参数组合最高产量的90% >,略高于CTL (Y结束= 89.64% (88.04;90.90])。农民之间的不平等略低于CTL结果(Yineq= 5.68% (4.97;6.34])。事实上,随着许可度的增加,在农民土地上进食的动物越来越少,因此对产量的影响更低,农民的产量接近最大值。最高的产量达到最大值,最低的产量不断增加,减少了不平等。
结果背后的机制
对于ATI,当监测到种群数量超过允许范围时,大多数灭绝事件发生,管理者相应地降低了扑杀限制的水平,以使种群减少到目标水平。在接下来的时间步骤中,当监视人口在允许范围内时,就会出现问题,因为这会导致管理人员保持策略不变。随后,农民继续以低成本进行扑杀,在下一个时间步将种群推向灭绝(图2,ATI面板)。因此,目标周围的允许范围越大,这种情况发生的可能性越大,从而解释了为什么消光频率和偏离目标的偏差随着允许度值的增加而增加。管理者的这种误传经常发生在消光频率非常高的ATI参数区域(图1),在消光前的时间步上,总体偏离目标的情况在管理者允许的范围内(附录2,图A2.4)。因此,在这里避免种群灭绝的最有效的策略是在每一个时间步中无条件地进行干预,以稍微降低农民的最终产量为代价。
轨迹策略
保护的结果
当应用TRJ时,对于高达80%的许可度值,消光频率和偏离目标的偏差至少与CTL一样接近于0,而管理者没有高达40%的时间进行干预(图1;附录3,无花果。A3.1和A3.2)。预算奖金值对结果要么没有影响,要么影响很小。几种组合的消光频率均在0.1以下,有时甚至为0fext= 0.15 (0.08;0.22] CTL。在40和50%的许可范围内(附录3,图A3.2),奖金数额的影响略强,奖金值在20和50%之间导致种群比CTL更接近目标(dT= -24.90% (-33.78;-16.26])。我们选择50%的参数区域作为管理者初始预算敏感性的实验,以测试当对管理者施加更强的预算约束时,这种弱效应是否会放大。
农业的结果
在使用TRJ时,无论许可度和预算奖金值如何,农民的最终产量都接近于CTL策略的最大景观生产能力(附录3,图A3.3)。同样,无论许可和预算奖金值如何,农民之间的产量不平等与CTL一样低(附录3,图A3.4)。
结果背后的机制
罕见的CTL灭绝事件似乎发生在种群数量超过目标值,管理者降低了过多的限制水平,或者农民碰巧扑杀了超出预期的数量,导致种群数量减少到无法弥补的情况下(图2,CTL面板)。TRJ策略可能避免了这种不精确性,它为管理者提供了在种群处于上限允许范围时不进行干预的可能性,并使种群接近目标(图2,TRJ面板)。预算奖金数额的缺失很可能是由于管理者的初始预算通常足以有效地确保人口维持和农民的产量,根据我们的初始参数值,没有由于奖金而留下改进的空间。因此,TRJ获得了与CTL类似的良好管理结果,而管理者无需在每个时间步骤进行干预,也无需考虑从等待期获得的收益。
对经理初步预算的敏感性
保护的结果
随着管理者初始预算的减少,灭绝频率增加,最终种群规模减少到目标以下(图3)B米= 800 b.u.,消光频率从0.71 [0.61;0.80]没有预算奖金,到0.07 [0.02;0.12]获得30%的奖金B米(图3),在相同的初始预算下,明显接近于零(fext= 0.76 (0.67;0.83])。在较高的奖金下,消光频率在0.3-0.6之间再次增加,虽然仍有较高的消光风险,但仍低于CTL。在目标距离上也有同样的趋势,从-78.4%上升TN(84.9;没有预算奖金,为-11.4% [-21.4;-2.1]同样是30%的奖金B米(附录4,图A4.1);CTL为-83.7% [-88.7;-78.0;](附录4,图A4.1)。
农业的结果
随着管理者初始预算的减少,农民的最终产量增加,农民之间的不平等减少(B米),因为它们对灭绝风险和种群规模都有积极的影响(附录4,图。A4.3和A4.4)。在B米= 800 b.u.面积时,最终产量在85% ~ 100%(绝种频率最高)之间,且不随红利量的变化而发生显著变化。加上30%的显著提高了保护效果的红利,最终产量为89.20% [87.47;90.76]而不是97.18% [96.14;99]同一个管理者的预算(以非常高的灭绝风险为代价)使用CTL策略。不平等为5.94% [5.23;6.68]而不是2.11% [1.65;2.6],仍然相对较低。
结果背后的机制
对于使预算奖金最大化的管理者初始预算值,其对灭绝风险的负作用和对种群规模的正作用(B米= 800 b.u),如果管理者在每一个时间步骤都进行干预,或使用TRJ但没有从等待期中得到任何好处,则当种群下降到过低的种群规模时,就会发生灭绝。由于扑杀成本总是过低,无法有效地降低农民的扑杀率(图4,CTL),因此,仅用最初的预算来纠正种群轨迹对管理者来说是一个挑战。在这种情况下,如果管理者从之前的等待期积累了预算奖金,那么一旦人口数量达到或预计达到管理者允许的范围,他们就有足够的权力对农民实施更高的限制。中间的奖金数额确保了当后者发生时,种群可以增加到更接近经理的目标(图4,TRJ)。因此,在对管理者的预算约束较强的情况下,TRJ似乎比CTL更有效。在这种情况下,预算奖金对降低灭绝风险至关重要,同时为农民保持高而公平的产量,并允许管理者节省20 - 30%的干预措施(附录4,图A4.2)。
讨论
研究总结
当对社会-生态系统中的保护冲突进行自适应管理时,我们对干预和等待之间动态交替的策略建模发现,当基于对系统响应的预测做出干预决策时,管理结果比仅基于最新的监测结果更好。通过基于预测的决策,保护和农业结果至少与无条件干预一样好,同时允许管理者节省管理资源,避免不必要的、潜在有害的干预。当低预算限制了管理者有效管理保护冲突的能力时,当干预最为关键时,等待期间累积的收益就会得到应用,保护结果会得到极大改善,而对农民的产量和权益影响甚微。自然,等待策略的主要风险是在需要干预时决定等待,或在等待更可取时决定干预。应该避免只根据当前的监测进行干预,因为当种群密度在急剧增加或减少的允许范围内监测时,管理人员可能错误地认为该政策是适当的,而实际上,保持相同的政策再次运行可能导致灭绝或临界产量损失。相反,基于人口轨迹的干预还包括不准确地预测人口密度在允许范围内的风险,导致管理者等待,而政策不足以协调保护和农业目标。然而,当使用适当的允许范围时,产量损失或种群下降的后果是可逆的。
预算和监控在等待策略效率中的重要性
我们的轨迹策略比无条件干预的优越性取决于管理者的预算。当预算足够高时,无论预算奖金多少,轨迹策略的结果至少与无条件更新一样好。这表明,当监测到的人口在允许的范围内,并预计将保持在该范围内(即,接近目标振荡)时,干预措施的作用较小。因为最初的预算足以满足满意的管理,在等待期间获得的收益与轨迹策略不能进一步改善管理结果。这是相关的,因为在保护方面,人力、财力和时间资源是有限的,在分配这些资源给案例方面存在不断的竞争(huhey等人2003年,McDonald-Madden等人2008年,Jachowski和Kesler 2009年,Ruiz-Miranda等人2020年)。人们也越来越认识到,在同一地理区域内,不同物种会在不同时间以不同方式影响人类生计,在管理中应考虑到这一点(Pozo et al. 2020, 2021)。因此,对一场冲突的干预可以在一段时间内成为优先事项,然后在另一场冲突需要更紧急的干预时被置于优先地位。因此,在资金充足的情况下,在等待期间未使用的资源可以分配给其他可能资金不足和/或更紧迫的情况,并提高整体保护效益(Wu等人,2020年)。因此,我们的轨迹策略可以帮助动态分配管理资源到在给定实例中最需要它们的案例中。
当有限的预算使管理更具挑战性时,不使用轨迹策略干预时节省的资源可以产生足够的效益来弥补资源的缺乏。我们强调,基于种群轨迹的预测是管理者降低干预时机误判风险的一种手段;这里改善管理的是更好地利用在等待期间积累的好处。这一结果支持了之前Iacona等人(2017)的建模结果,即国家公园管理者没有足够的预算将每一个濒危鸟类物种都置于保护之下,但可以通过等待和节省资金来逐步增强其货币权力来最大化成功。重要的是,只有在不需要时不撤销或重新分配未使用的管理资源时,这才可能实现。Ruiz-Miranda等人(2020)对保护中的退出策略进行了综述,发现在适应性管理中,当目标达到时退出资金是非常罕见的(但应该更多地考虑和仔细规划)。本研究建议,如果管理资源不受限,则应重新分配在等待期间节省的预算;如果管理资源受限,则应投资于改进未来干预措施。
为了分离不同的时间策略对管理质量的影响,我们假设管理者对种群大小有完全的了解。但现实世界的监测涉及不确定性,这在保护的成功中发挥了重要作用(Bunnefeld et al. 2011, Nuno et al. 2013)。监测不确定性会导致估计种群密度的误差,从而导致判断是否需要干预的误差。这将降低无条件干预和弹道策略的效率,但后者可能受到更大的影响,因为误差将影响监测和弹道预测,因此减少了优于无条件干预的优势。事实上,弹道战略的有效性可能依赖于更定期和更准确的监测,这可能并不总是可能的或负担得起。测试观察准确性或成本对管理质量的影响超出了本研究的范围,但在应用计时策略时,这是需要考虑的一个重要方面(McDonald-Madden et al. 2011, Milner-Gulland 2011, Wu et al. 2020)。
因为我们关注的是管理策略而不是控制措施,为了简单和易于模型解释,我们将农民的选择限制在扑杀上。我们没有对间接措施进行建模,比如在土地使用保护冲突管理中广泛使用的围栏(Nyhus 2016, Pooley et al. 2017),因为这些措施是相当永久性的建筑,并不总是适合我们适应性管理过程的定期变化和更新。尽管如此,未来的模型可能有用地考虑到人口管理的一系列备选办法。
自适应管理的建模新特性
正在进行的6th快速变化的气候下的大规模灭绝(Ceballos et al. 2017)以及农业和野生动物保护之间的土地利用冲突对粮食安全的后果往往使保护管理者处于紧急状态(Du Toit 2010)。我们的研究结果表明,行动的紧迫性不应意味着系统的、无条件的干预,而应强调获取信息以明智地选择干预方式和时间的重要性。与渔业管理中的is -fish (Mahévas和Pelletier 2004)或FLR (Kell et al. 2007)等软件一样,这里开发的方法可以为管理者制定政策提供信息。用瑞典养殖鹤与普通鹤之间的冲突的经验数据参数化GMSE,此前已允许评估补贴水平,以最好地平衡扑杀和恐吓,以维持鸟类数量和农民的收入(Nilsson等人,2021年)。同样,我们的模型的目标参数化可以为管理者提供信息,以决定他们应该有多宽容,以及他们应该从我们的战略在保护和土地使用者的目标和管理资源配置效率方面的有用的等待期中期望获得多少收益。
我们模型基于个体的特性和GMSE框架的模块化解释了围绕种群动态和利益相关者个人决策的几个不确定性来源。我们的机制模型模拟了具有内在人口不确定性(生活事件实现的个体间变异性)和地理不确定性(动物的移动是随机的;uchmakovsky和Grimm 1996, Stillman et al. 2015)。未来的工作还可能包括对环境不确定性的明确建模,可能以随机极端事件的形式影响人口动态和农民产量。目前,即使种群动态是不确定的,如果空间分布有时会导致一个农民的土地上的动物数量多于另一个农民,我们的结果仍然是稳健的。Rakotonarivo等人(2020,2021)表明,在利益相关者群体之间的保护行动的成本和收益平衡中,感知到的公平性在土地使用者选择支持保护战略的倾向中发挥着重要作用。然而,在建模结果中很少考虑保护冲突中的公平问题。例如,Wam等人(2016)在其管理模式中使用了不同利益相关者群体之间的货币权益衡量标准,以平衡北方森林中的伐木、牲畜放牧和狩猎活动。我们的方法还通过系统地面对人口动态和农民的产量来控制利益相关者之间的公平。此外,我们使用了一个新的指标参数来衡量利益相关者之间的权益,通过衡量我们策略的成功与最低和最高农民产量之间的差异。 Among-stakeholder equity, to our knowledge, has not been modeled before in conservation conflicts, and modeling stakeholders individually like the present study offers a direct measure of equity among members of the same group, thus allowing its monitoring as an important outcome of management.
缺乏动态的涉众行为建模被认为是保护失败的主要原因(Schlüter等,2012)。之前的研究通过使用博弈论建模决策来解决这一问题(Colyvan等人,2011年,Glynasti等人,2018年)。然而,当应用于管理决策时,游戏理论框架可能有局限性,包括固定的行为规则,有限的行动集(例如,合作或背叛),以及假设玩家是完全理性的,知道对他们来说最好的选择(Myerson 1997)。在这个模型中,我们使用进化算法(一种人工智能的形式)让管理者和农民做出决策,我们表明,当选项面板对于博弈论问题来说太大时,它提供了一种启发式的方法来找到实际的解决方案(Hamblin 2012)。我们将进化算法与基于个体的方法相结合,并为每个利益相关者独立建模决策,并在连续的可能行动中考虑次优选择的可能性(另见Kamra等人2018年,Cusack等人2020年,Nilsson等人2021年)。在我们的实验中模拟这些不同的不确定性来源,使我们能够得出这样的结论:即使管理者并不总是制定最有效的政策,如果农民并不总是按照预期行事,我们提出的策略也是相关的。
结论
我们使用了一个不确定性-鲁棒建模工具,比较了在保护和农业目标方面的等待策略和无条件干预的管理质量,并根据保护冲突的案例讨论了使用哪种策略。我们提出了一种管理人员动态交替干预和等待的策略,根据人口监测的信息。当干预或等待的决定是基于对人口轨迹的预测时,我们的策略可以更好、更公平地管理保护冲突,特别是在预算有限的情况下。通过在不需要干预时节省时间、精力和/或金钱,它还可以确保更有效地利用管理资源。
致谢
A.B.由IAPETUS NPIF拨款资助,资助代码NE/R012253/1,由NERC支持;a.b.d由勒弗休姆信托基金早期职业奖学金资助;J.M.和N.B.是由欧洲研究理事会根据欧盟H2020/ERC拨款协议no 679651 (ConFooBio)资助的。
数据可用性
支持这项研究结果的数据/代码可以在GitHub上https://github.com/AdrianBach/EcologyAndSocietyArticle-data-code公开获取。该研究获得了斯特灵大学的伦理批准(批准号EC 2018_19 342)。
文献引用
班布里奇,i . 2017。苏格兰鹅的管理概况。中46:224 - 230。https://doi.org/10.1007/s13280-016-0883-5
贝尔,D. M., A. Ozgul和G. Cozzi。2017。在统一的社会生态适宜性模型中结合人类接受度和生境适宜性:以瑞士狼为例。应用生态学报54(6):1919-1929。https://doi.org/10.1111/1365-2664.12880
Bunnefeld, N, c.t.t. Ewdards, A. Atickem, F. Hailu,和E. J. Milner-Gulland。2013.鼓励对战利品狩猎的监督和遵守。保护生物学27(6):1344 - 1354。https://doi.org/10.1111/cobi.12120
Bunnefeld, N, E. Hoshino,和E. J. Milner-Gulland。2011.管理策略评估:保护的有力工具?生态与进化动态26(9):441-447。https://doi.org/10.1016/j.tree.2011.05.003
塞巴洛斯,G. P. R. Ehrlich和R. Dirzo. 2017。正在进行的第六次生物大灭绝以脊椎动物数量的损失和下降为标志。中国科学院学报114(30):E6089-E6096。https://doi.org/10.1073/pnas.1704949114
M. Colyvan, J. Justus和H. M. Regan. 2011。保护的游戏。生物保护144(4):1246 - 1253。https://doi.org/10.1016/j.biocon.2010.10.028
库萨克,J. J.杜西,J.敏德曼,I. L.琼斯,R. A.波佐,O. S. Rakotonarivo, S. Redpath和N.邦内菲尔德,2020。综合冲突、游说和服从来预测自然资源使用的可持续性。生态与社会25(2):13。https://doi.org/10.5751/ES-11552-250213
迪安吉利斯,D. L.和V.格林,2014。四十年后生态学的个体基础模型。F1000Prime代表6(39)。https://doi.org/10.12703/P6-39
杜塞,A. B., J. J.库萨克,I. L.琼斯,J. Minderman, E. B.尼尔森,R. A.波佐,O. S. Rakotonarivo, B. Van Moorter, N. Bunnefeld. 2018。GMSE:通用管理战略评估的R包。生态学与进化方法9(12):2396-2401。https://doi.org/10.1111/2041-210X.13091
杜·托伊特,2010。生物多样性保护中的规模考虑。动物保护13:229 - 236。https://doi.org/10.1111/j.1469-1795.2010.00355.x
弗雷克塞尔、J. M.帕克、K.麦肯、E. J.索尔伯格和b . e。年代æ其他。2010.资源管理周期和收获野生动物种群的可持续性。科学》328(5980):903 - 906。https://doi.org/10.1126/science.1185802
Game, E. T. Meijaard, D. Sheil和E. McDonald-Madden。2013.在一个邪恶复杂的世界里保存;挑战和解决方案。保护信7(3):271 - 277。https://doi.org/10.1111/conl.12050
Glynatsi, n.e, V. Knight和T. E. Lee. 2018。犀牛角贬值的演化博弈模型。生态建模389:33-40。https://doi.org/10.1016/j.ecolmodel.2018.10.003
Gregory, R., D. Ohlson, J. Arvai. 2006。解构适应性管理:应用于环境管理的标准。生态学应用16(6):2411 - 2425。https://doi.org/10.1890/1051 - 0761 (2006) 016 (2411: DAMCFA) 2.0.CO; 2
格林,v . 1999。生态学中基于个体的建模十年:我们学到了什么,未来又能学到什么?生态建模115(2 - 3):129 - 148。https://doi.org/10.1016/s0304 - 3800 (98) 00188 - 4
汉布林,s . 2012。遗传算法在生态学和进化中的实际应用。生态学与进化方法4(2):184-194。https://doi.org/10.1111/2041-210x.12000
希克斯、L. L.、W. O.沃格尔、D. R.赫特、R. J.厄尔利和H. C.斯塔宾斯。2009。第21章-梅子溪中央瀑布栖息地保护规划和北方斑点猫头鹰模型。J. J.米尔斯波和F. R.汤普森编辑的561-592页。大型景观野生动物保护规划模型。学术,剑桥,马萨诸塞州,美国。https://doi.org/10.1016/b978-0-12-373631-4.00021-6
休伊,K. F. D., R.卡伦和E.莫兰,2003。在保护管理中,将经济学纳入优先事项的确定和评估。保护生物学17(1):93 - 103。https://doi.org/10.1046/j.1523-1739.2003.01317.x
Iacona, g.d., h.p. Possingham和M. Bode, 2017。等待可能是一种最佳的保护策略,即使在危机学科中也是如此。中国科学院学报114(39):10497-10502。https://doi.org/10.1073/pnas.1702111114
杰考斯基,D. S.和D. C.凯斯勒,2009。允许物种灭绝:我们应该让物种消失吗?生态学与进化动态24(4):180。https://doi.org/10.1016/j.tree.2008.11.006
Kamra, N., U. Gupta, F. Fang, Y. Liu, M. Tambe. 2018。基于神经网络的连续空间安全博弈策略学习。人工智能第三十二届AAAI会议论文集。2018年2月2日至2月7日,美国路易斯安那州新奥尔良。https://feifanginfo.files.wordpress.com/2017/11/aaai_2018_optgradfp.pdf
卡斯瓦米拉,A., S.拉塞尔,M.麦克吉本。2007.野生动物对坦桑尼亚东北部家庭粮食安全和收入的影响。野生动物的人类维度12(6):391-404。https://doi.org/10.1080/10871200701670003
基思,D. A., T. G.马丁,E.麦克唐纳-马登和C.沃尔特斯,2011。生物多样性保护的不确定性与适应性管理。生物保护144(4):1175 - 1178。https://doi.org/10.1016/j.biocon.2010.11.022
凯尔,L. T., I.莫斯基拉,P.格罗斯让,j . m。弗罗曼汀,D.加西亚,R.希拉里,E.贾迪姆,S.马德尔,M. A.帕斯托斯,J. J.普斯,F.斯科特,R. D.斯科特。2007。FLR:一个用于评估和发展管理战略的开源框架。海洋学报64(4):640-646。https://doi.org/10.1093/icesjms/fsm012
Mahévas, S.和D. Pelletier. 2004。用于评估管理措施对渔业动态的影响的通用和空间显式模拟工具is - fish。生态建模171(1 - 2):65 - 84。https://doi.org/10.1016/j.ecolmodel.2003.04.001
曼利,B. F. J. 2007。生物学中的随机化、自举和蒙特卡罗方法。第三版。查普曼和霍尔/CRC,伦敦,英国。
梅森,T. H. E, A.基恩,S. M.雷德帕特和N.邦内菲尔德。2017。环境保护冲突的变化:苏格兰的鹅和农业。应用生态学报55(2):651-662。https://doi.org/10.1111/1365-2664.12969
梅森,T. H. E, C. R. J.波拉德,D.奇马拉康达,A. M.格雷罗,C.科尔-史密斯,S. A. G.米尔赫拉斯,M.罗伯茨,P. R.纳法克,和N.邦内菲尔德。2018。恶冲突:用恶问题思维对保护冲突进行整体管理。保护信11 (6):e12460。https://doi.org/10.1111/conl.12460
麦克唐纳-马登,P. W. J.巴克斯特和H. P.波辛厄姆,2008。在有限的资金和知识下,为保护环境做出强有力的决策。应用生态学报45(6):1630-1638。https://doi.org/10.1111/j.1365-2664.2008.01553.x
麦克唐纳-马登,E. I. Chadès, M. A.麦卡锡,M.林基,和H. P.波辛厄姆。2011。将保护资源分配给物种存活不确定的地区。生态学应用21(3):844 - 858。https://doi.org/10.1890/09-2075.1
Milner-Gulland, 2011。综合渔业方法和家庭实用模式,以改进资源管理。中国科学院学报108(4):1741-1746。https://doi.org/10.1073/pnas.1010533108
迈尔森,R. B. 1997。博弈论:冲突分析。马萨诸塞州剑桥,哈佛大学出版社,美国马萨诸塞州剑桥。https://doi.org/10.2307/j.ctvjsf522
尼科尔,S., K.沃德,D.斯特拉特福德,K. D.乔尼克和我Chadès。2018.最好地利用专家的估计来确定监测和管理行动的优先次序:淡水案例研究。环境管理学报215:294-304。https://doi.org/10.1016/j.jenvman.2018.03.068
Nilsson, L., N. Bunnefeld, J. Minderman和A. B. Duthie. 2021。利益相关者赋权对鹤种群数量和农业生产的影响。生态建模440:109396。https://doi.org/10.1016/j.ecolmodel.2020.109396
努诺,A., N. Bunnefeld,和E. J. Milner-Gulland。2013.将观测结果与现实相匹配:使用模拟模型来改善塞伦盖蒂不确定性下的监测。应用生态学报50(2):488-498。https://doi.org/10.1111/1365-2664.12051
尼胡斯,2016。人类与野生动物的冲突与共存。环境与资源年度评论41:143-171。https://doi.org/10.1146/annurev-environ-110615-085634
Pérez, P., A. Ruiz-Herrera和A. M. San Luis. 2020。干扰倾向人群的管理指南:干预时间的重要性。理论生物学杂志486:110075。https://doi.org/10.1016/j.jtbi.2019.110075
普利、S、M.巴鲁阿、W.贝纳特、A.迪克曼、G.霍尔姆斯、J.洛里默、A. J.洛夫里奇、D. W.麦克唐纳、G.马文、S.雷德帕斯、C.西勒罗-祖比利、A.齐默尔曼和E. J.米尔纳-古尔兰。2017.当前和未来改善人类捕食者关系的方法的跨学科综述。保护生物学31(3):513 - 523。https://doi.org/10.1111/cobi.12859
波佐,R. A, J. J.库萨克,P. Acebes, J. E. Malo, J. Traba, E. C. Iranzo, Z. maurice - trainor, J. Minderman, N. Bunnefeld, S. Radic-Schilling, C. A. Moraga, R. Arriagada和P. Corti. 2021。协调家畜生产和野生草食动物保护:挑战与机遇。生态与进化动态36(8):750-761。https://doi.org/10.1016/j.tree.2021.05.002
波佐,R. A, E. G. LeFlore, A. B. Duthie, N. Bunnefeld, I. L. Jones, J. Minderman, O. S. Rakotonarivo和J. J. Cusack. 2020。野生动物对当地社区生计影响的多物种评估。保护生物学35(1):297 - 306。https://doi.org/10.1111/cobi.13565
Rakotonarivo, o.s., a.r. Bell, K. Abernethy, J. Minderman, A. B. Duthie, S. Redpath, A. Keane, H. Travers, S. Bourgeois, l - l。穆卡格尼,J. J.库萨克,I. L.琼斯,R. A.波佐和N.邦内菲尔德,2021。基于激励的手段和社会公平在保护冲突干预中的作用。生态与社会26(2):8。https://doi.org/10.5751/ES-12306-260208
Rakotonarivo, O. S, I. L. Jones, A. Bell, A. B. Duthie, J. J. Cusack, J. Minderman, J. Hogan, I. Hodgson和N. Bunnefeld. 2020。保护冲突干预的实验证据:财政支付、社区信任和公平态度的重要性。人与自然3(1):162-175。https://doi.org/10.1002/pan3.10155
雷德帕斯,S. M., R. J. Gutiérrez, K. A.伍德,J. C.杨。2015。保护中的冲突走向解决方案。剑桥大学出版社,英国剑桥。https://doi.org/10.1017/cbo9781139084574
雷德帕斯,s.m., J.杨,A.伊夫利,W. M.亚当斯,W. J.萨瑟兰,A.怀特豪斯,A.阿玛尔,R. A.兰伯特,J. D. C.林内尔,A.瓦特,R. J. Gutiérrez。2013.理解和管理保护冲突。生态与进化动态28(2):100-109。https://doi.org/10.1016/j.tree.2012.08.021
理查德森,S. A. C.米尔,D.戴维斯,D.詹姆,A. I.沃德,2020年。为控制入侵性、非本地哺乳动物和其他人类与野生动物的冲突而进行的适应性野生动物管理的系统综述。哺乳动物回顾50(2):147 - 156。https://doi.org/10.1111/mam.12182
Ruiz-Miranda, c.r, L. I. Vilchis和R. R. Swaisgood, 2020年。野生动物保护的退出策略:为什么它们是罕见的,为什么每个机构都需要一个。生态环境科学进展18(4):203-210。https://doi.org/10.1002/fee.2163
Schlüter, M, R. R. J.麦卡利斯特,R.阿灵豪斯,N.邦内菲尔德,K.爱森纳克,F. Hölker, E. J.米尔纳-古兰,B. Müller, E.尼克尔森,M.夸斯,M. Stöven。2012.环境管理的新视野:社会-生态系统耦合模型综述。自然资源建模25(1):219-272。https://doi.org/10.1111/j.1939-7445.2011.00108.x
C.西姆斯和D.芬诺夫,2013。什么时候”等着瞧吧”对入侵物种采取措施是否合理?资源与能源经济35(3):235-255。https://doi.org/10.1016/j.reseneeco.2013.02.001
A. D. M.史密斯,K. J.塞恩斯伯里和R. A.史蒂文斯,1999。实施有效的渔业管理制度-管理战略评价和澳大利亚伙伴办法。海洋学报56(6):967-979。https://doi.org/10.1006/jmsc.1999.0540
斯蒂尔曼,R. A., S. F. Railsback, J.吉斯克,U.伯格,V.格林,2015。在一个变化的世界中做预测:基于个体的生态学的好处。生物科学65(2):140 - 150。https://doi.org/10.1093/biosci/biu192
乌奇马耶斯基,J.和V.格林,1996。生态学中基于个体的建模:是什么造成了差异?生态与进化动态11(10):437-441。https://doi.org/10.1016/0169 - 5347 (96) 20091 - 6
范维尔根,B. W.和H. C.比格斯,2011。对南非大草原保护区适应性生态系统管理的关键评估。生物保护144(4):1179 - 1187。https://doi.org/10.1016/j.biocon.2010.05.006
沃尔特斯,1986。可再生资源的适应性管理。麦克米伦,纽约,纽约,美国。
Wam, H. K, N. Bunnefeld, N. Clarke, O. Hofstad. 2016。生态系统服务利益冲突:货币和非货币措施之间权衡的多标准建模和间接评估。生态系统服务22 (B): 280 - 288。https://doi.org/10.1016/j.ecoser.2016.10.003
威廉,B. K., F. A.约翰逊,K.威尔金斯,1996。水禽收获的不确定性与适应性管理。野生动物管理学报60(2):223-232。https://doi.org/10.2307/3802220
吴,学术界。,A. J. Dodd, C. E. Hauser, and M. A. McCarthy. 2020. Reallocating budgets among ongoing and emerging conservation projects. Conservation Biology 35(3):955-966.https://doi.org/10.1111/cobi.13585

图1

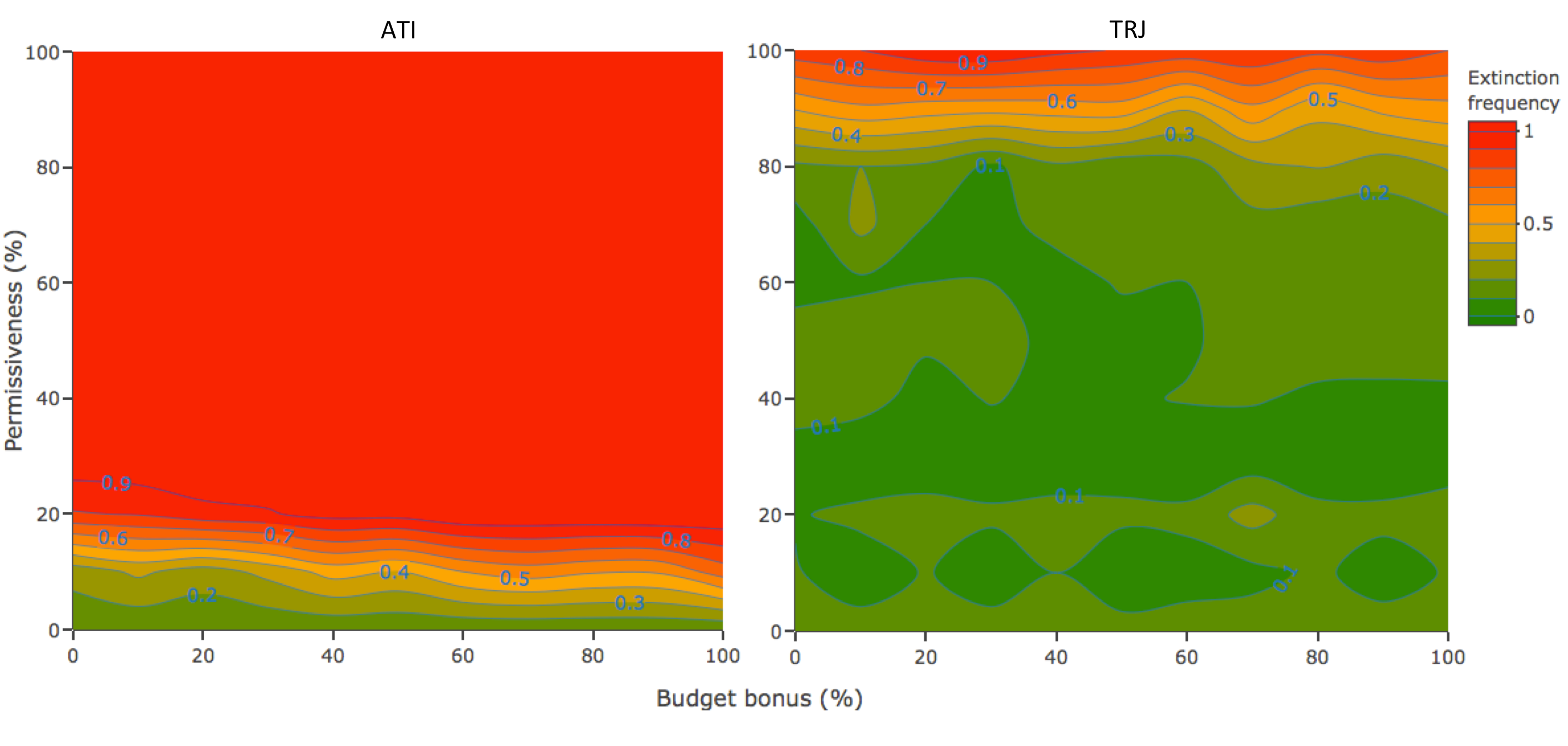
图1.灭绝的频率(fext)根据许可(PT)及预算奖金(Bb)组合在一个以个体为基础的模型中,模拟在保护冲突条件下的种群管理。越绿,灭绝的风险越低。乐队是由PT= 0和相应的Bb值fext由控制策略(CTL)得到。具有自适应时间干预策略(ATI;左面板),没有组合PT而且Bb参数导致低afext作为对照策略(CTL;0.15 (0.08;0.22] 95% CI),多数情况下种群灭绝几乎是确定无疑的,具有弱正效应Bb不管允许的范围大小。利用轨迹策略(TRJ;右侧),大多数地区的绿色和CTL一样或更绿fext值,意味着TRJ在灭绝风险方面的表现至少与CTL一样好。的影响Bb在fext虚弱到缺席。
图2
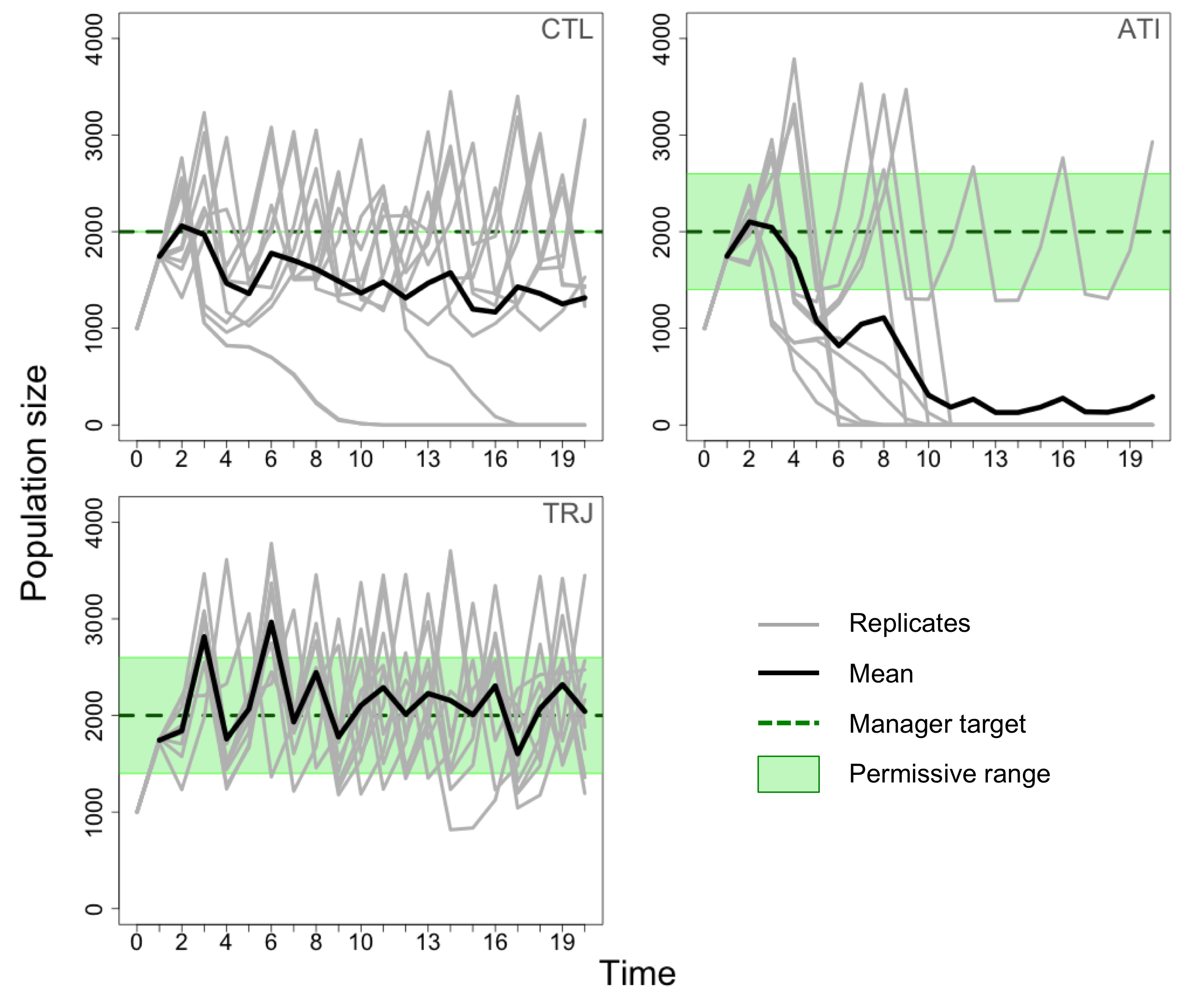
图2.使用基于个体的模型模拟种群在保护冲突条件下的适应性管理,10个模拟重复的平均种群大小。
左上:当管理者无条件干预时(控制策略,CTL)。当种群数量远远低于目标数量时,就会发生灭绝(TN),以便管理器能够通过增加限制进行纠正。
右上:应用自适应时机干预策略(ATI;放纵(PT) = 30%,预算奖金(Bb) = 10%)。大多数物种灭绝发生在种群规模超过允许范围的时候,然后在接下来的时间步骤进行监测。因此,管理人员没有更新政策,允许农民以低成本继续扑杀,经常在接下来的时间步骤中导致种群灭绝。注意:在没有导致灭绝的复制中,种群在减少的过程中从未被监测到允许范围内,这使得管理员更新成本,以更好的时机控制情况。
左下:应用轨迹策略(TRJ;TN= 30%,Bb= 0%)。TRJ策略避免了一些灭绝事件。
图3
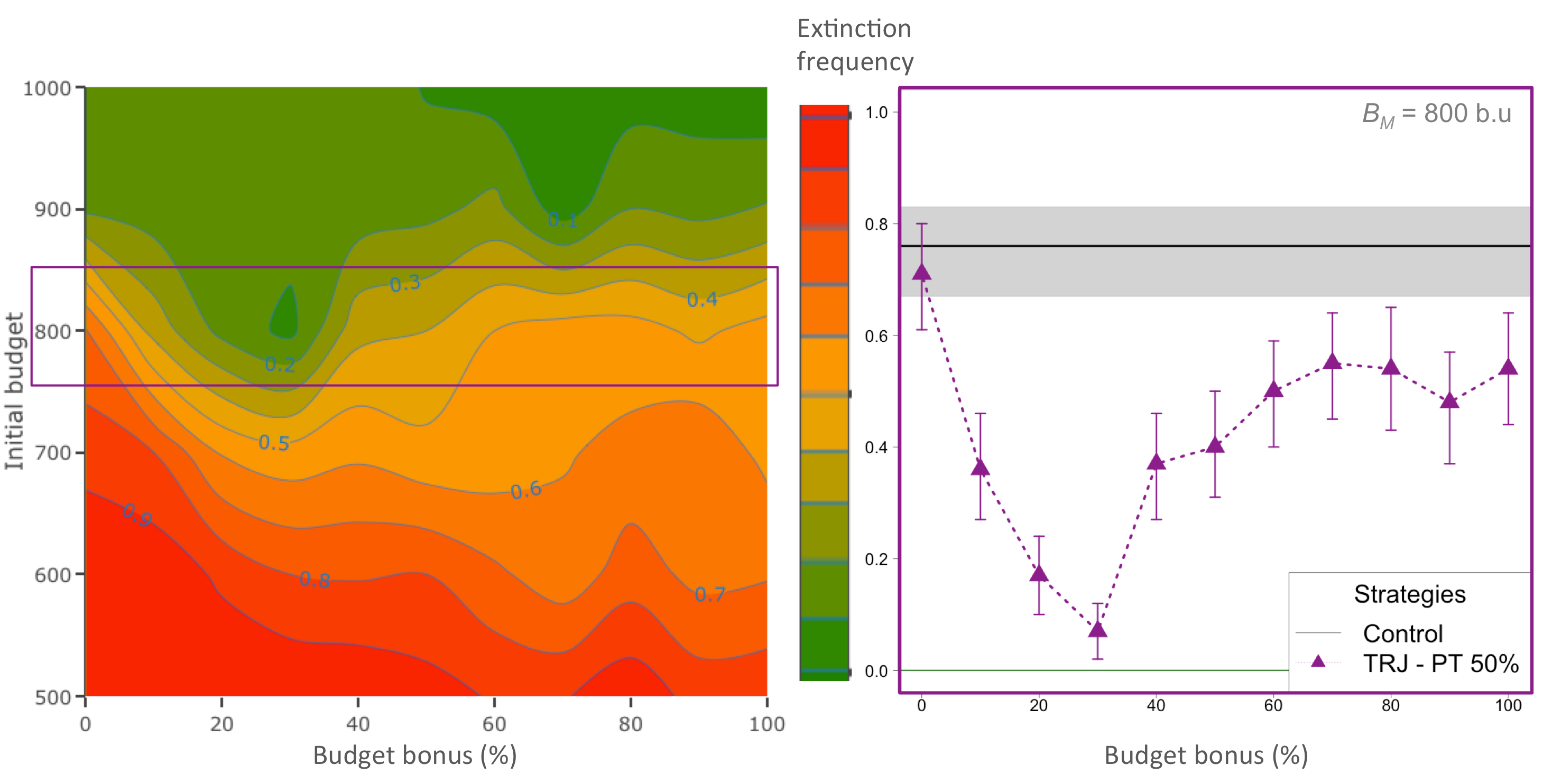
图3.应用轨迹策略时的消光频率(TRJ);放纵(PT) = 50%,根据经理的初步预算(B米)及预算奖金金额(Bb)在一个基于个体的模型中,模拟了在保护冲突条件下种群的适应性管理。绿色越绿,消光频率越低。为B米= 800 b.u.(紫色正方形,在右边的面板上的细节),沿着增加的方向形成一个坑Bb值,意思是低到中间值Bb明显降低了灭绝的风险。误差条显示95%的引导置信区间。黑线是fext控制策略为相同的初始预算和灰色阴影区域的95%置信区间。
图4
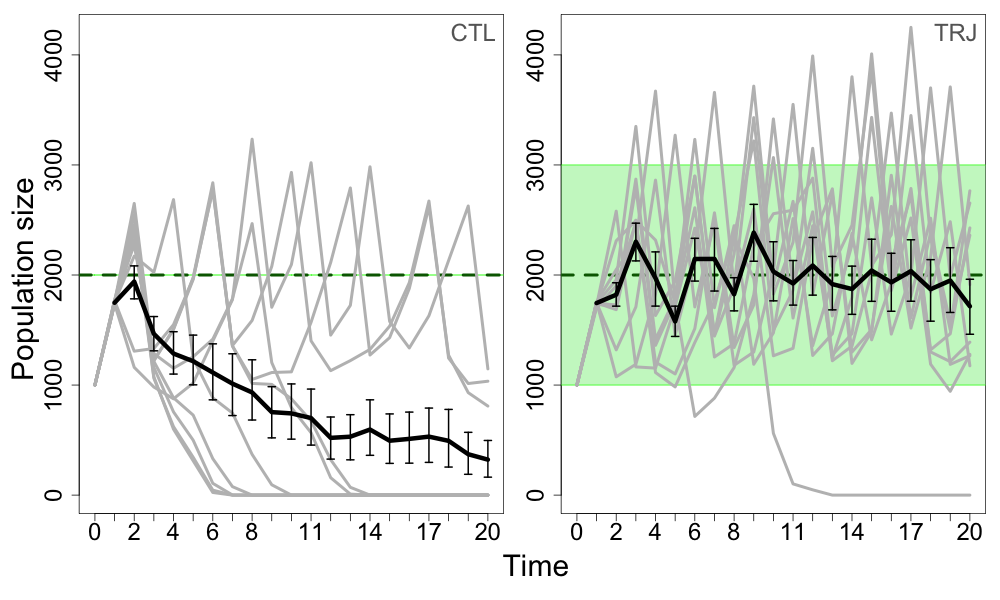
图4.在10个重复(灰色细线)上绘制了一个基于个体的模型,模拟了在保护冲突条件下种群的适应性管理,初始预算为800 b.u。绿色虚线显示了管理者的目标(TN),绿色区域为允许范围TN±PT.
左面板:应用控制策略(CTL)时。当种群数量在两个连续时间步骤中远远低于管理者的目标(绿色虚线)时,就会发生灭绝,而管理者只能通过最初的预算进行调整。
右面板:当应用轨迹策略(TRJ;PT= 50%,Bb= 30%)。由于等待期间积累的收益,经理能够将扑杀成本提高到足够高的水平,使农民的扑杀率保持在一个可持续的价值。导致灭绝的复制是由于对时间步骤10的种群规模的严重错误预测造成的,导致管理员在需要干预时等待。