






以下是引用这篇文章的既定格式:
蒂托,R., N.萨利纳斯,E. G.科西奥,T. E.波萨·埃斯皮诺萨,J. G. Muñiz, S. Aragón, A.尼娜,R.罗曼-奎斯塔。2022.秘鲁的次生林:与其他森林砍伐后过渡相比,生态系统服务的提供有所不同。生态与社会27(3):12。摘要
由于人类的直接影响,热带森林正在迅速转变,许多森林被砍伐的地区正在通过自然或人类协助的再生恢复为森林。这种情况为执行森林管理战略以实现环境目标同时促进社会发展和促进当地生计提供了一个机会之窗。然而,成功的森林管理政策取决于我们如何评估环境后果以及这些再生森林提供的生态系统服务的价值。在这里,我们回顾了已发表的文献,综合了秘鲁沿海、安第斯和亚马逊地区三种类型的森林过渡所提供的生态系统服务:自然再生次生林、农林复合系统和人工林。然后,我们将讨论这些重新生长的森林作为基于自然的解决方案的潜力,这些解决方案有助于制定促进森林可持续利用和保护的政策。我们的文献分析表明,森林转型在抵消碳排放、为生物多样性提供栖息地和调节水文服务方面提供了重要的服务。然而,生态系统服务的数量和重要性因森林过渡类型而异。次生林提供多种服务,是减轻气候和生物多样性危机的一种低成本、即时和高度有效的战略,并最终为社会提供重要的生态系统服务,例如供水。相反,外来树种种植对水调节服务有负面影响。我们强调次生林在土地管理方面的潜力,为多种综合环境倡议提供支持。 This framework can guide policy decisions to choose appropriate options on forest transition types most suitable to achieve specific end goals at local and regional scales, considering both ecosystem services and disservices to avoid trade-offs in which the achievement of one goal is detrimental to another.介绍
热带成熟森林储存了大量的碳并拥有高度的生物多样性,但由于直接的人类影响和不断变化的环境条件,它们正在迅速转变(Hansen等人,2013年;Colorado Zuluaga和Rodewald, 2015年;粮农组织,2020年)。秘鲁森林退化的情况与其他国家没有太大的不同。秘鲁国家森林和野生动物局(SERFOR)在其最近关于森林砍伐的报告中透露,2017年至2019年期间,秘鲁亚马逊雨林的森林损失率平均为每年128069公顷,与前10年相比有所增加(https://geo.serfor.gob.pe/visor/).同样,其他研究表明,安第斯山脉和沿海地区许多地区的森林扰动率不断上升(Aide等人2019年,粮农组织2020年)。森林损失的主要原因是为了满足对粮食和其他基本产品日益增长的需求而扩大农业和牧场。此外,采金导致的森林砍伐也造成了很大程度的森林损失(Tovar等人,2013年,Caballero Espejo等人,2018年,Csillik和Asner 2020年)。在短期内,这种土地使用变化带来的直接和可观察的经济效益超过了森林生态效益的损失,而森林生态效益往往是人类感知不到的(D 'Almeida等人,2007年,Giam 2017年,Moomaw等人,2019年)。
许多因木材开采、农业或牧场而砍伐森林的地区正在通过自然或人类协助的再生(即植树;查兹登等人,2020年)。这些”新森林”或”重新造林”通常是在一段与社会经济变化相一致的森林砍伐时期之后,以不断增加的趋势恢复其森林覆盖的森林被称为”森林转换”(Wilson et al. 2017, MacDonald and McKenney 2020)。在许多地区,废弃毁林土地上的自然再生森林(即次生林)正在日益扩大,并可能在不久的将来成为热带森林的主要特征(Poorter et al. 2016, Wilson et al. 2017)。最近,据估计次生林占热带地区剩余森林的一半(McGee等,2020年)。据报道,秘鲁亚马逊地区约260 - 800万公顷的土地是次生植被(MINAM 2015年,Smith等人,2021年)。已发表的报告显示,次生林通常会迅速积累地上生物量,从而以比成熟林更高的速度吸收二氧化碳(Asner等人2010年,Poorter等人2016年,Chazdon等人2016年)。除了碳封存,次生林还拥有高度的生物多样性,并产生关键的供应生态系统服务,如水、木材或食物(Poorter等人,2016年,Jones等人,2019年)。尽管有这些和其他重要的好处,次生林在生态和经济方面仍然被低估,因此在森林管理政策中基本上被忽视。
除了次生林,热带景观也越来越多地被农林复合系统和人工林所占据(Horgan 2009, Socolar等,2019)。尽管这些系统不是自然再生的森林,但许多研究主张,这些由人类管理的树木覆盖区域具有提供重要服务和效益的潜力(Ehrenbergerová等,2016年,Zavala等,2018年,Jezeer等,2019年)。但是,它们在生态上彼此不同,也不同于原始的土地覆盖(即成熟的森林;Wilson et al. 2017),以及它们最终提供的环境后果以及生态系统服务的价值仍然知之甚少(Barbier et al. 2010, Wilson et al. 2017, MacDonald和McKenney 2020)。例如,某些类型的林场(如外来树种林场)的扩张提供了重要的供应和支持服务,但也可能降低其他服务,如与水和土壤相关的服务(Raboin和Posner, 2012, Vallet等人,2016,Bonnesoeur等人,2019)。当森林的真正利益不为人知时,也会导致森林政策的不明确。从这个意义上说,同时分析森林覆盖类型、结构、功能和服务的变化可以为设计适当的土地管理战略提供有价值的信息(MacDonald和McKenney 2020)。因此,估计不同森林类型所提供的生态和社会效益是必要的第一步。
鉴于人们日益认识到气候和土地覆盖变化的影响,提出了一些国际和国家倡议,以保护完整的森林,减少森林砍伐,并通过自然或辅助森林再生恢复退化的土地(Shukla等人,2019年)。森林和景观恢复议程寻求通过树木生长从大气中去除二氧化碳来限制全球气候变化,这是目前正在采用的此类举措的一个例子(Shukla等人2019年,Cbeplay竞技hausson等人2020年,Soto-Navarro等人2020年)。其中一项活动是波恩挑战,这是一项到2030年恢复3.5亿公顷土地的国际承诺(Holl和Brancalion 2020;https://www.bonnchallenge.org/).根据20x20倡议(2019年布宜诺斯艾利斯宣言:https://initiative20x20.org/news/buenos-aires-declaration-restoration),秘鲁已承诺恢复320万公顷土地,其中200万公顷用于商业树木种植,其余120万公顷用于不同退化形式(过度放牧、盐碱化、水侵蚀、土壤污染和土壤压实)的土地;Román等2018,Cerrón等2019)。然而,这些策略面临着重大挑战。造林和再造林在执行和长期监测方面都要花费大量的时间和金钱。事实上,长期监测往往不到位,造林和再造林的短期成功并不能转化为持久和有效的森林恢复(Cerrón等,2017年,2019年,SERFOR 2018年)。相比之下,自然再生次生林可能是优化多种生态系统服务提供的更合适的方法,因为这些森林是在环境条件允许的地区恢复的,绕过了可能影响在退化土地上建立人工造林和再造林的供水和气候变化问题(Chazdon和Guariguata 2016年)。确定哪种森林过渡类型能够在短期内提供多种和有效的服务,对于寻找缓解和适应气候变化的有效工具以及支持综合、可持续的土地利用管理极其重要(Chazdon和Guariguata, 2018年)。beplay竞技
为了帮助改善秘鲁自然再生森林的决策和管理,我们研究了有关不同的以树木为基础的恢复退化土地的方法的现有文献。我们只关注森林转型,因为在当前气候变化、生物多样性丧失和荒漠化的挑战下,与其他农业选择相比,森林转型提供的多种服务被认为是双赢的局面。beplay竞技在这里,我们分析了秘鲁三个主要地区(沿海、安第斯和亚马逊)的三种不同的森林过渡(次生林、农用林业系统和人工林)及其提供的三种关键生态系统服务(碳封存和储存、生物多样性栖息地和水调节)。我们还提供了关于成熟原生森林所提供的生态系统服务的信息,以提供与这些森林过渡所提供的生态系统服务相比较的参考状态。更具体地说,我们解决了以下关键问题:我们可以从次生林和其他森林砍伐后的过渡中期待什么生态系统服务?这些系统在每个地区的相对重要性是什么?这些信息如何帮助设计有效的社会和环境政策?
研究背景和数据来源
研究背景
秘鲁拥有丰富多样的生态区,在世界上104个生态区中总共有84个,以及28种不同的气候(Escobal和Torero 2003年,MINAM 2014年)。秘鲁地貌的异质生境维持了高度的生物多样性,为社会提供了重要的服务。秘鲁国民经济的约22%与生物多样性和生物多样性产品贸易有关,约为2亿美元(MINAM 2014年)。与此同时,秘鲁被认为是世界上最易受气候变化影响的国家,并受到土地利用变化的高压(MINAM 2015年,2016年,Gobierno del Perú 202beplay竞技0)。因此,由于其不同的地理特征和依赖自然的经济活动,秘鲁面临着来自全球变化影响的严重挑战。在这方面,森林将在保护生物多样性和减缓气候变化及其相关影响方面发挥关键作用。beplay竞技
亚马逊森林:秘鲁的植被覆盖1.03亿公顷,其中7200万公顷为森林,大部分位于海拔500米以下的亚马逊低地(MINAM 2015年)(图1A)。该地区是秘鲁人口最少的地区,但其森林采采量最大。截至2017年,整个亚马逊盆地17%的原始森林面积已被砍伐(Bullock等人,2020年),秘鲁亚马逊地区6.9%的原始森林面积已被砍伐(Smith等人,2021年)。2000-2020年期间,秘鲁低地亚马逊森林的平均年森林损失为128,764公顷/年(粮农组织2020年)。亚马逊地区的农业和金矿开采正以前所未有的速度扩张,这些活动是秘鲁森林砍伐的主要导火索(Caballero Espejo等人,2018年,Manoli等人,2018年,Csillik和Asner, 2020年)。例如,在过去20年里,秘鲁亚马逊东北部的油棕种植园的扩张大幅增加,从2000年的1.5万公顷增加到2019年的10.8万公顷,导致2.8 Pg的碳排放(Escobedo Grandez 2021年)。2007-2013年农业扩张导致的森林砍伐中,油棕种植园约占11% (Vijay等人,2018年)。相比之下,金矿开采造成1.12 Tg/年的碳排放(Csillik和Asner 2020年),其影响在秘鲁,尤其是在Madre de Dios (Caballero Espejo等人,2018年,Álvarez-Berríos等人,2021年)。
安第斯山脉的森林:安第斯地区(占该国国土面积的27%)由一条从北到南跨越该地区的纵向山脉组成(图1A)。从干燥的西坡海拔2000米,湿润的东坡海拔500米,到海拔6000米冰川的高寒地区。2011年,安第斯山脉的森林面积为1220万公顷,约占全国森林总面积的21% (MINAM 2015, 2016)。这些森林在提供水文服务和支持生物多样性方面具有不成比例的重要性,但快速的土地利用变化对生物多样性和生态系统服务的可用性构成了日益严重的威胁。因此,咖啡、可可和茶园(图1B-D)以及大火、造林和外来树种再造林正在增加安第斯自然土地覆盖的压力(Tovar等人2013年,粮农组织2016年,Oliveras等人2018年)。2018年,荫咖啡农林业约占安第斯山脉总面积的5%(608,332公顷)(粮农组织2020年)。到2012年,有100万公顷安第斯非林区(天然草地和退化土地)被造林(粮农组织,2016年)。这些种植园大多专注于异域风情桉树而且松果体物种的快速生长和短期内的经济盈利能力(粮农组织2016年,Cerrón等人2017年,SERFOR 2018年)。例如,在卡哈马卡地区,桉树而且松果体在1987年至2007年期间,人工林取代天然草地的速度以12.3%/年的速度增长(Tovar et al. 2013)。除了人类的影响外,安第斯山脉的气候正在迅速变化,气温上升导致冰川融化,改变了天气模式,以及相关的长期干旱(Vuille et al. 2008, 2018)。
干燥的沿海地区和季节性落叶林:该地区占秘鲁总面积的11.7%,是一条从太平洋延伸到海拔2000米的干燥的西安第斯斜坡的狭长纵向地带(SERFOR 2019;图1 a)。尽管水资源有限,但该地区居住着约55%的人口(MINAM 2016),秘鲁首都利马位于西部沿海沙漠。尽管很少有研究对拉丁美洲和秘鲁的干森林砍伐率进行估计,但在秘鲁沿海森林中,森林砍伐确实是一个严重的问题(Whaley et al. 2010, Pécastaing和Chávez 2020)。例如,在皮乌拉地区,据估计每年损失1.5万至4万公顷干森林(Pécastaing和Chávez 2020)。城市对柴火和木炭的需求,加上农业和畜牧业的扩张,是干燥森林保护的主要威胁(Whaley等人,2010年,Bennett-Curry等人,2013年,SERFOR 2019年)。这些人类活动加剧了气候变率增强的影响,例如与厄尔Niño-Southern振荡相关的事件(Pécastaing和Chávez 2020),在秘鲁北部表现为沿海降水和洪水增加(El Niño-Costero),在南部表现为严重干旱(Bourrel et al. 2015)。
数据源
我们对谷歌Scholar数据库中截至2020年9月的英语和西班牙语出版物进行了搜索,并对已发表的同行评审文章、硕士论文、博士论文和研究报告进行了审查和分析,针对我们选定的三种森林转型:次生林、人工林和农林复合体系的生态系统服务主题进行了审查和分析。在人工林类别中,我们包括商业、单一栽培和异国情调的人工林。在农林复合系统类别中,我们包括了间作系统、树阴多年生系统、防风林和活栅栏系统。考虑到当前和未来的环境变化条件,我们将重点放在对自然和人类福祉至关重要的生态系统服务上。因此,我们回顾了至少评估了以下三个生态系统服务之一的出版物:碳储量和封存、生物多样性栖息地和水调节。我们使用了不同关键词的组合(”二次森林”,”第二个地面森林”,”农林复合经营”,”树种植园”,”自然再生”等),以找出与我们研究兴趣有关的文献。尽管最初专注于同行评议的期刊文章,我们扩展了我们的文献搜索到”灰色文献”(即硕士或博士论文、研究报告)识别所有与评审主题相关的可用数据。由于在搜索中检索到大量的论文,我们根据标题和摘要进行了第一轮筛选,以选择与我们的目标最相关的文章。最初共有163篇摘要被确定为符合纳入标准,但进一步筛选的结果是共76篇论文被纳入了本综述。只考虑了在秘鲁进行的研究。
结果与讨论
森林生态系统服务
在森林过渡理论(即从砍伐森林转向增加森林覆盖)中,森林通常被简单地定义为森林覆盖,并假设森林覆盖的增加可以改善生态系统服务(Barbier等人2010年,Garcia等人2020年)。然而,森林提供独特的服务,其质量和数量取决于其类型、特征和环境条件。出现这种情况是因为不同的森林恢复路径和驱动因素加上不同的生态特征、分布和轨迹导致了不同的生态系统功能和森林服务(Barbier等人2010,Vallet等人2017,Wilson等人2017,Garcia等人2020,MacDonald和McKenney 2020)。
通过文献分析,我们发现经验证据表明,再生森林为广泛的野生动物分类提供了重要的碳汇和栖息地(方框1-3,图2和3)。森林覆盖恢复对水文也有积极或消极的影响,这取决于森林类型和区域的具体特征(方框1-3)。正如预期的那样,次生林、农用林业系统和人工林提供的生态系统服务的数量和价值因森林类型和区域而异。
亚马逊森林生态系统服务
亚马逊森林是现存最大的陆地碳储量(例如,Saatchi et al. 2007, 2011, Asner et al. 2010)。特别是秘鲁低地亚马逊森林,其地上生物量有潜力储存150毫克/公顷的碳(Asner等,2014年)。最近的一项研究报告称,在整个秘鲁亚马逊地区,有260万公顷次生林,这种森林恢复导致了73.8 Tg的碳积累(Smith等,2021年)。
我们的文献综述显示,亚马逊次生林是在保护生物多样性的同时缓解气候变暖的有希望的途径(框1,图2A)。次生林更高的生产力意味着次生林的生长速度和碳吸收比成熟林快11倍(Poorter et al. 2016)。然而,最近的研究表明,这种增强的生长受到反复干旱的严重影响,次生林的吸收可能被高估了(Smith et al. 2020)。研究表明,与其他森林路径相比,次生林的碳储量恢复速度快于人工林和农林复合系统(框1,图2A)。秘鲁亚马逊地区30年生次生林储存了成熟森林价值的50-60% (Málaga等,2021年),50年生次生林储存的碳量与成熟森林相当(Chuquizuta等,2016;框1,图2A)。尽管农林复合系统和人工林储存了大量的碳,但与成熟森林转换相比,它们仍然造成了很高比例的碳损失,特别是当人工林是快速生长、低木材密度的外来物种时(框1,图2A)。这些发现支持了之前研究(Wilson等人,2017年,Lewis等人,2019年)的主张。
除了碳封存潜力外,次生林还拥有大量的生物多样性(框1,图2B)。例如,30年至50年的次生林拥有附近成熟森林中80% - 100%的鸟类、哺乳动物和爬行动物物种(Whitworth et al. 2016),是大型动物特别重要的栖息地(Tapirus terrestris,大的灵长类动物,Priodontes马克西姆斯)和一些受威胁的物种(例如,彪马concolor;加文·2004)。相比之下,农林系统和人工林的结构比次生林简单得多,为较小的物种提供了栖息地(框1,图2B)。农用林业系统中蜣螂和白蚁物种的恢复比次生林高,农用林业系统和人工林中昆虫物种恢复的比例也很高(框1,图2B)。人工林的某些组成和结构特征可以促进某些物种的发生。例如,在巴西亚马逊地区,大量的成熟森林蝙蝠、表栖蛛形动物、蜥蜴和屎壳郎物种在具有原生灌木下层植被的外来树种种植园地区被发现,类似于次生林中出现的那些物种(Barlow et al. 2007)。生物多样性保护的价值不仅在于维持物种和遗传多样性,还在于它们为人类带来的利益和服务。因此,在自然再生地区出现的许多野生树木和动物也为当地居民提供了重要的食物来源和额外收入(Gavin 2004, 2007, Fitts et al. 2020)。
在这里,我们总结了秘鲁亚马逊地区次生林和毁林后森林过渡(农林复合系统和人工林)提供的三种关键生态系统服务的主要发现。完整的研究清单及其碳储量值和物种分布见附录1和附录2。
碳储量
成熟的森林
报告的亚马逊低地地上碳储量在107.6 - 335.1毫克/公顷之间(Martel和Cairampoma 2012, Sosa Castillo 2016)。总碳储量在155.6-632.4毫克/公顷之间(Barbarán 1998, Martel和Cairampoma 2012)。
次生林
研究报告表明,地上碳储量和总碳储量都随着林龄的增加而增加(图2)。例如,早期(1-5年生)、幼龄(15年生)和老龄(50年生)次生林地上碳储量分别为2.4-42.1 Mg/ha、131 - 184.4 Mg/ha和~282.8 Mg/ha (Alegre et al. 2003, Chuquizuta et al. 2016)。同样,早期次生林、年轻次生林和老次生林的总碳储量分别为40.5-95.8、153.3-239.1和396.8 Mg/ha (ICRAF 1998, Alegre et al. 2003, Chuquizuta et al. 2016)。平均而言,30年次生林储存成熟林价值的50-60% (Málaga等,2021年),而50年次生林储存的碳量与成熟林相当(Chuquizuta等,2016年)。
农林复合经营系统
亚马逊低地的许多农林系统都是如此Theobroma可可或Coffea阿拉比卡树荫下有两到六种树木。农用林业中最常见的树种是Inga edulis, Guazuna crinite, Calycophyllum spruceanum, C. spruceanum, Cedrela odorata, Mariosousa willardiana, Cajanus cajan, Schizolobium amazonicum而且银合欢leucocephala。地上碳储量在1年的农用林业系统中为2.9-4.4 Mg/ha,在20年的农用林业系统中为27-32.4 Mg/ha,而总碳含量在1年和10年的农用林业系统中分别为33.4-37.2 - 168.9 Mg/ha (Concha et al. 2007, Villogas Ventura 2013, Timoteo et al. 2016)。然而,农林复合系统的碳储量取决于树种,因此这些值可能有所不同(附录1)。
树种植园
人工林地上碳储量在1年生时为9.5 Mg/haGuazuma crinita(Baltazar Peña 2011) 30年至66.6 Mg/ha橡胶树取代巴西橡胶树种植园(Alegre et al. 2003)。30年总碳储量为152.3-152.6 Mg/ha橡胶树取代巴西橡胶树种植园(ICRAF 1998, Alegre et al. 2003)。
而不是森林,6岁和30岁Bactris gasipaes据报道,人工林的总生物量中碳含量分别为72.2和90.9 Mg/ha (Chuquizuta et al. 2016, Cuellar Bautista和Salazar Hinostroza 2016)。
维持生物多样性的生境
次生林
几项研究表明,自然再生森林,特别是老次生林的生物多样性很高;30 - 50年的次生林拥有附近成熟森林中73-95%的两栖动物、81-98%的鸟类、79-81%的哺乳动物和88-110%的爬行动物物种(Whitworth et al. 2016)。相比之下,年轻的次生林(9-15岁)含有74-83%的屎壳螂,36.8%的白蚁,以及周围成熟森林中发现的47%的树种(Horgan 2009, Culot et al. 2011, Gonzalez et al. 2014, Vebrova et al. 2014, Duran-Bautista et al. 2020;图2B,附录2)。
农林复合经营系统
在亚马逊低地,农用林业系统包含高度多样性的蜣螂(87%)和昆虫(71-79%),但很少有树木(23-24%)、白蚁(47%)和鸟类(14-16%)物种(Horgan 2009, Lojka等人2011,Vebrova等人2014,Perry等人2016,Aerts等人2017,Duran-Bautista等人2020;图2B,附录2)。
树种植园
人工林的生物多样性低于成熟林、次生林和农林复合系统。与成熟森林相比,林树人工林的昆虫和屎壳郎种类分别为66.7%和30-39% (Horgan 2009, Lojka et al. 2011, Aerts et al. 2017;图2B,附录2)。
虽然不是真正的森林种植园,但油棕(Elaeis guineensis)种植园中包含了附近成熟森林中发现的60.5%的白蚁和42.6%的鸟类(Srinivas和Koh 2016, Duran-Bautista等人,2020)。
水文服务
入渗和土壤湿度:尽管森林砍伐可能会减少当地降水,导致土壤干燥和蒸散减少(Marengo 2006, D 'Almeida et al. 2007),但本地树种的自然更新改善了降雨和渗透,增加了有效水分(Brandon 2014)。自然再生可以(在5-10年内)迅速逆转对水调节的影响(Hölscher等,2005年)。在可可复合林业系统中,遮荫树产生的植物凋落物的积累(印加sp。大叶蕨、云杉蕨、麻叶蕨、而且牡荆pseudolia)允许更大的雨水入渗,防止土壤水分流失,增加土壤保水能力(Arévalo-Gardini et al. 2015)。但农林复合土壤植物有效水分(10.6 ~ 11.7%)显著低于30年次生林土壤(12.3 ~ 13.4%);Arévalo-Gardini et al. 2015)。
水流调节及水质:热带成熟森林提供了高度的水渗透,很少侵蚀或地表径流,并通过防止一些沉积和侵蚀改善水质(Marengo 2006, D 'Almeida et al. 2007, Brandon 2014)。亚马逊成熟森林比人类改造的植被类型(人工林或农业;Bruijnzeel 2004)。蒸散发是潮湿亚马逊地区的一个关键过程,占当地降水输入总量的28% (Ellison等,2012年)。相比之下,幼树人工林的蒸散量高(类似于成熟森林),但入渗较低,减少了地下水补给(Brandon 2014)。同样,人工林取代成熟森林导致蒸发损失增加,导致年流量减少(Scott et al. 2005)。
油棕种植园的扩张在过去几十年里蓬勃发展,导致生态系统退化(Butler和Laurance 2009年,Srinivas和Koh 2016年)。油棕幼林显著降低了生态系统的蒸散量(无林下植被时为- 40%,有地被覆盖时为- 20%)和渗透率,并显著增加了径流(无林下植被时高达40%;Manoli等人。2018)。水流的这些变化导致当地气候变热和变干(即产量的变化)。然而,成熟人工林(8年生)的初级生产力更高,蒸腾的水更多(比被取代的森林多7.7%),从而减少了水径流(Manoli等人,2018年)。
亚马逊低地地区的森林砍伐通过减少当地降水和增加干旱强度对当地水循环造成严重影响(Marengo 2006, D 'Almeida et al. 2007)。森林再生可以通过在当地和区域水文过程中提供重要服务来扭转这些影响,改善入渗、降雨和湿度(专栏1)。研究表明,亚马逊次生林在多个尺度上改善了淡水的质量、产量和输送,从地下水到河流和降雨。农林复合作物还能改善雨水入渗,从而提高蓄水能力(方框1)。相比之下,人工林会产生负面影响,增加用水成本,减少入渗,降低地下水补给和年流量(方框1)。例如,人工林(例如,桉树、松果体、橡胶树)取代原始森林导致蒸发需求增加,导致年流量减少(Scott et al. 2005)。总的来说,我们强调了在这个对社会和环境具有重要生态系统服务的高生物多样性地区不断扩张的大规模单种商业种植园对水文的潜在有害后果(专栏1)。
安第斯森林生态系统服务
安第斯地区生物多样性异常丰富,是全球生物多样性的热点地区,具有高度的地方性(Myers et al. 2000)。它在土壤中提供了大量的碳储存,特别是在积水的土壤中(Román-Cuesta et al. 2011),它是两个安第斯斜坡的基本区域水供应者:干燥的西太平洋海岸(Beresford-Jones 2004, Whaley et al. 2010)和东海岸,其亚马逊河的源头是安第斯溪流(Clark et al. 2014, Doornbos 2015)。
秘鲁热带安第斯森林的生物量表现出与多变的环境和地形条件有关的高度变异性。然而,研究表明,三种过渡的地上生物量在森林覆盖恢复的头几十年表现出相对相似的值,人工林的储量最高(图3框2)。例如,约10年次生林、农用林业系统和人工林的地上生物量平均含碳43、54和72 Mg/ha,约占成熟林储量的20、27和33%(框2,图3)。图3).在安第斯山脉高地区,云山森林的生长速度很低,大约30年的次生林在其地上生物量中只储存了32%的成熟森林碳(Oliveras et al. 2018)。相比之下,这些山地云林可以有相对较高的土壤碳积累率,由于苔藓的丰富(特别是泥炭藓)储存水分并防止凋落物分解(Oliveras等人,2018年)。安第斯次生林的土壤恢复比其他人为管理的森林系统更快(Oliveras等,2018年,Walentowski等,2018年)。
在这里,我们总结了秘鲁安第斯地区次生林和毁林后森林过渡(农林复合系统和人工林)提供的三种关键生态系统服务的主要发现。完整的研究清单及其碳储量值和物种分布见附录1和附录2。
碳储量
成熟的森林
秘鲁山地森林的地上碳储量为52.4-485.3 Mg/ha (Lapeyre et al. 2004, Oliveras et al. 2018),总碳储量为154.3-398.8 Mg/ha (Chuquizuta te al. 2016, Oliveras et al. 2018)。
在高海拔的安第斯山脉,森林斑块以Polylepis而且Escallonia物种储存的碳量低于其他山地森林,地上生物量为4.8-40.1毫克/公顷,总碳储量为23.8-148.7毫克/公顷(Cuellar Bautista和Salazar Hinostroza 2016年,Gurmendi Porras和Orihuela Izaguirre 2019年)。
二次森林
早期(1 ~ 5年生)次生林地上碳储量为26.1 Mg/ha,幼龄(~10年生)和老龄(~30年生)次生林地上碳储量大致相当,分别为43.2 Mg/ha和36.6 ~ 42.3 Mg/ha。平均而言,与成熟林相比,约30年的次生林仅储存32%的地上生物量碳(Oliveras等,2018,Aragón等,2021)。早、幼、老次生林总碳储量随林龄增加而增加(分别为116.6、161.1和295.3 Mg/ha碳)。
农林复合经营系统
咖啡农林复合林地地上碳储量印加、松果体、或桉树碳储量在19.3 ~ 62 Mg/ha之间,总碳储量在119.9 ~ 177.5 Mg/ha之间。碳储量的变化是由于用作荫庇的年龄和物种(Lapeyre et al. 2004, Ehrenbergerová et al. 2016)。
树种植园
林地地上碳储量随树龄增加,5- 10年从30.1 Mg/ha增加到72-72.7 Mg/ha蓝桉或松果体放射虫纲种植园。同样,这些种植园的总碳储量分别为129和136.2-142.3毫克/公顷(Gamarra Ramos 2001年,Cuellar Bautista和Salazar Hinostroza 2016年,Bernachea Jesus 2019年)。然而,Raboin和Posner(2012)报告说,28岁的年轻人松果体杉木人工林仅储存35.7毫克/公顷的碳。
维持生物多样性的生境
次生林
热带山地云雾林火灾后10年次生林包含未烧毁地区75% - 93%的植物物种,而老烧毁地区(10-28年)的多样性比附近成熟森林多72% (Oliveras et al. 2014)。同样,次生林的蜣螂种类比周围成熟森林多66.7% (Vélez Quesquén和Saavedra Chávez 2019)。相比之下,老次生林支持成熟森林中36-105.4%的鸟类物种(Colorado Zuluaga and Rodewald 2015, Hosner et al. 2015;附录2)。
农林复合经营系统
荫荫咖啡农林业比附近的成熟森林拥有更多的鸟类物种多样性(120.3%),但农林业系统中的大多数物种是通才(Colorado Zuluaga和Rodewald 2015;附录2)。
树种植园
与成熟森林相比,桉树而且赤杨皮(桤木)人工林拥有79-120.8%的灌木和70.2-84.2%的土壤大型动物物种(de Valença et al. 2017)。没有找到其他分类单元的信息。
水文服务
产水量和贮水量:次生林缓冲峰值流量和在土壤中储存水分的能力略高于外来人工林(估计优势比:2.22 vs. 2.37;Bonnesoeur等人,2019)。相比之下,农林复合系统(荫凉咖啡)和人工林(前牧场)的水分产量分别比成熟森林低11%和14% (Quintero等,2009年)。其他研究也显示了人工林的类似水文模式。重新造林种植奇异树木(松,蓝桉,柏树)超过1%的集水区如果取代放牧或天然草地,则会导致水量分别减少20%和40% (Bonnesoeur et al. 2019)。同样,在安第斯高原草原上最古老的松树种植园比天然草地土壤少保留63%的水分(Farley et al. 2004),并减少约50%或平均每年242毫米的水产量(Buytaert et al. 2007)。相比之下,在农林复合林地和林分林地,水分调节只受到轻微影响(桉树viminalis而且石莲子spinosa)在安第斯山脉西坡(Villar Cabeza et al. 2014)。
入渗和土壤湿度:在退化土壤上造林14 - 20年,入渗率提高了8倍,但比成熟的原生森林低3倍(Bonnesoeur等,2019年)。土壤含水量Polylepis次生林(在以前的马铃薯田)的水分含量略低于附近的天然草地(50-74%),而20年松树种植园的土壤水分含量显著低于天然草地(13-22%)(Harden et al. 2013)。
水流的规定:用松树造林桉树在西安第斯地区,由于水的使用增加,该物种减少了9-11%的地表径流和6-8%的流量(Krois和Schulte 2013年)。水平均每天流入桉树绿化区域比邻近的自然集水区低4-10倍(Ochoa-Tocachi et al. 2016, Ochoa-Tocachi 2019)。
空气湿度截取:在农林业中,与次生林和人工林类似,无论树木密度如何,相对冠层截水量占总降雨量的25% (Bonnesoeur等人,2019年)。
水质:松林对水质的影响最小(van Dijk和Keenan, 2007)。
与亚马逊地区的次生林类似,安第斯山脉的次生林拥有高度多样性的鸟类、树木和屎壳虫(附录2框2)。在热带山地云林中,火灾发生的频率相对较高,并对生物多样性造成严重影响(Young和León 2007, Oliveras et al. 2014),但在被烧毁的地区可以发生快速和旺盛的新生(Román-Cuesta et al. 2011, Oliveras et al. 2014, 2018)。允许物种提前快速恢复(Oliveras等,2014;在农林复合系统和人工林中,管理人员经常采用技术来控制树木的密度和多样性,并控制影响其他物种的害虫和杂草。此外,一些外来树种种植园(例如,桉树、广泛种植在安第斯山脉)释放化感化合物,阻止本地物种的形成。这可能是农用林业系统和人工林包含较少或主要是多面手昆虫种类的一些原因(框2)。
我们的文献综述显示,安第斯山脉的森林对水文服务有积极或消极的影响,这取决于森林类型(框2)。天然森林的更新和本地物种的再造林改善了水的供应和调节服务,而外来树种的人工林对水文调节有消极的影响。特别是当它们取代了天然草地(专栏2)。这些外来的人工林减少了河流流量和地下水补给,因为它们消耗大量的水,并通过蒸腾作用释放出来。这种情况意味着下游用户的可用水量减少,特别是在旱季(方框2)。
沿海森林生态系统服务
尽管关于秘鲁沿海地区的干森林及其生态系统服务的信息很少,但众所周知,干森林是当地社区极其重要的生存和收入来源(由森林提供的木材、柴火和食物)Prosopis物种)。沿海干燥森林受到森林砍伐的威胁,尽管它们的树木稀疏(在大多数沿海地区的覆盖率小于30%)和占主导地位的algarrobo物种(Prosopis pallida;利昂·卡塞雷斯2019)。尽管干燥森林的碳储量低于安第斯和亚马逊森林(框3),但这一生态系统中的原生植被更新尤其明显Prosopis物种在提供关键的生态系统服务,如供水服务和生物多样性栖息地方面发挥着重要作用。例如,干燥次生林支持较高的地方性,这导致树木和哺乳动物(如食虫蝙蝠)的多样性相对较高(框3)。Prosopis树木可使土壤湿度增加28%,从深层土壤层吸收水分,并从大气湿度中捕获大量水分(框3)。由于在漫长的旱季,海岸几乎不存在降雨,植被通过雾捕获和凝结提供了每年大量的水供应(Whaley et al. 2010)。沿海干燥森林植物已经进化出适应能力(框3)来在这些环境条件下生存。气孔调节等适应性调节水分消耗,使物种能够忍受干旱胁迫(Time et al. 2018);小的和木质化的叶分枝,或”brachyblats”,捕捉大气湿度(Whaley et al. 2010);而且”逆液压升降机”提供了在夜间吸收大气水分的能力(Bereford-Jones 2004)。最近的一项研究表明,这些森林在减少人类对厄尔尼诺的脆弱性Niño方面具有多么重要的作用,因为干燥森林覆盖的丧失放大了干旱的负面影响并增加了当地温度(Pécastaing和Chávez 2020)。
在这里,我们总结了秘鲁沿海地区次生林和森林砍伐后过渡(农林复合系统和人工林)提供的三种关键生态系统服务的主要发现。完整的研究清单及其碳储量值和物种分布见附录1和附录2。
碳储量
成熟的森林
据报道,沿海干林地上碳储量平均为27.6 Mg/ha,总碳储量平均为44.1 Mg/ha (Cuellar Bautista和Salazar Hinostroza 2016)。
次生林
25年次生林地上生物量碳储量为4.23 Mg/ha(相对于成熟林约15%),总碳储量为46.9 Mg/ha(与成熟林相似;查韦斯苏亚佐2018)。
树种植园
地上的碳存量Mangifera籼人工林的碳储量为8.2 Mg/ha,总碳储量为14.3 Mg/ha (Cuellar Bautista和Salazar Hinostroza 2016)。
农林复合经营系统
我们没有发现农林业系统碳储量的数据,这表明需要在该地区进行更多的研究。
利用激光雷达遥感和包括所有森林类型的地上碳含量估算表明,秘鲁沿海干燥森林平均储存1-8±2毫克/公顷的碳(Asner等,2014年)。
维持生物多样性的生境
二次森林
秘鲁沿海干燥的次生森林支持52.2-70%的树种(Rasal Sánchez et al. 2011, Delgado Paredes et al. 2020)和附近成熟森林中发现的166.7%的食虫蝙蝠物种(Ruiz Romero 2015)。然而,与成熟森林相比,秘鲁西北部的次生林只包含22.7%的鸟类物种(León Cáceres 2019;附录2)。
农林复合经营系统
Gossypium而且穆萨在成熟干燥森林中发现的鸟类中,79.6%是农林复合系统的鸟类(Chávez-Villavicencio 2013)。
水文服务
在干旱的秘鲁沿海地区Prosopis物种是在水生态系统服务中发挥重要生态作用的最具代表性和最常见的物种。此功能由以下机制介导:
- 的深二形根系(可达60米)Prosopis从深层土壤中获取水分,并将部分水分与其密集的表层根系网络一起沉积,使上层土壤(< 35 cm)的水分增加28% (Beresford-Jones 2004, Whaley et al. 2010)。深层根系吸收的水分在夜间通过根系移动并释放到上层土壤剖面(称为“水力提升”)的机制在干旱生境生长的其他几种物种中得到了证实(Caldwell和Richards 1989年,Caldwell等人1998年,Time等人2018年)。在干旱的环境中,液压提升的水形成了一个缓冲供应,在白天为邻近的物种和提升植物本身平衡了水分压力(Beresford-Jones 2004)。
- 在近生芽胞体密集簇中的小小叶(最小的木质化叶枝)Prosopis适应于凝结大气湿度并将其沉淀到树冠下(Beresford-Jones 2004, Whaley et al. 2010)。例如,一个小的p . pallida一棵树(3米高,树冠4米高)每晚可吸收高达9升的水(Whaley et al. 2010)。Prosopis在夜间,它还通过叶子吸收大气中的水分,并将水分固定在土壤中(通过一种叫做”逆液压升降机”;Beresford-Jones 2004)。这一机制至关重要,因为秘鲁沿海地区的许多地区基本上不存在降水,而雾可以占到水流的50% (Brandon 2014年)。
- Prosopis是高度干旱地区有效用水的使用者。例如,p . juliflora每年用水量约为235毫米,仅占20%桉树camaldulensis相同条件下的用水量(Mahmood et al. 2001, Beresford-Jones 2004)。
总的来说,Prosopis生成”岛屿的湿度”在自然和人工改造的森林过渡中,可以提供重要的供水效益(Beresford-Jones 2004)。
总之,我们的综合研究表明,所有森林过渡类型都提供了有价值的服务。然而,重要的是要考虑所提供的生态系统服务的数量和重要性。尽管农林复合系统和人工林提供了碳储量和野生动物栖息地服务,但它们仍然导致生态系统服务损失的比例很高。此外,外来树种种植对水文服务有负面影响,从而减少了可用水。相比之下,我们发现次生林是一种更有效和直接的森林类型,可以优化各种生态系统服务的提供,我们将在下面进一步探讨。
次生林是基于自然的解决方案
次生林在减缓气候变化方面的作用beplay竞技
与人类足迹对地球生态系统的巨大影响相比,气候变化迄今为止所起的作用相对较小。beplay竞技然而,人类世及其人类活动的干预增加了气候模式的不确定性(例如,不断变化的降雨模式、极端温度波动、极端事件发生频率更高),使减缓和适应气候变化、保护生物多样性和保障人类福祉变得更加困难(Malhi等人,2020年)。beplay竞技必须立即采取雄心勃勃的减缓气候变化行动,以减轻社beplay竞技会和生态系统必须面对的影响的严重性。为了实现这一目标,以自然为基础的解决方案正在作为综合方法出现,可以帮助减少和保护人类免受气候变化影响,同时减缓全球变暖,支持生物多样性,并确保生态系统服务的提供(Griscom等人,2017年,Chausson等人,2020年,Seddon等人,2020年)。beplay竞技次生林在以自然为基础的解决方案中发挥着至关重要的作用,例如自然气候解决方案(与更成熟的森林相比,次生林的隔离率更高,具有缓解潜力)、绿色基础设施(次生林有助于防止侵蚀和减少风暴影响)和以生态系统为基础的适应方案(由于土壤入渗率更好,可以储存水和安第斯森林中的苔藓等蓄水选项)。
根据《联合国气候变化框架公约》(UNFCCC),秘鲁至少有两种直接方法可以将次生林纳入基于自然的解决方案。beplay竞技一种方法是将次生林的缓解潜力作为国家预定国家确定贡献的一个明确界定的组成部分。第二种方法是秘鲁承诺恢复120万公顷退化土地(在20x20倡议下承诺的320万公顷土地中)。2015年9月,秘鲁政府向《联合国气候变化框架公约》提交了第一份国家自主贡献文件,承诺到2030年将温室气体排放量减少30%,比预计的“照常发展”水平低(Gobierno del Perú 2016)。秘鲁在最近修订的重新提交的文件中显示了更大的雄心,承诺在未来十年减少40%的排放量(Gobierno del Perú 2020)。为实现这一承诺而作出的约70%的缓解努力考虑到改变土地利用和改善林业部门(包括造林和农业效率;Gobierno del Perú 2016, Gonzales-Zuñiga 2018, Gallice等2019)。然而,秘鲁政府仍然没有采用这一战略(气候行动跟踪2019年),也没有明确考虑次生林的潜力(Gobierno del Perú 2020)。因此,秘鲁目前的气候政策行动被称为”不足”不符合《巴黎协定》1.5°C的限制(气候行动跟踪2019年)。据估计,改善森林管理可使秘鲁实现目前有条件目标的32%(气候行动跟踪2019年)。目前,秘鲁缺乏关于次生林管理的国家政策。安第斯和沿海地区的次生林在研究、保护和可持续利用方面比亚马逊森林更被忽视。这种缺乏可能部分归因于秘鲁环境部,该部仍在寻找和量化全国各地次生林的碳储量和减缓潜力。我们强调,迫切需要作出更大努力,加快采取促进次生林管理、常生林保护、将原生林和再生林纳入生态系统服务付费范围的政策,以促进其持久性。将次生林包括在国家确定的贡献和倡议20x20承诺范围内的一个常见问题是,次生林只被视为碳。然而,正如这里和其他研究所显示的,这些森林提供的服务远不止碳(Lewis等人,2019年)。
秘鲁和拉丁美洲其他地区的缓解活动正在鼓励采用非本地单一树种的低生物多样性造林和再造林商业做法(Zamorano-Elgueta等人,2015年,Moomaw等人,2019年,Heilmayr等人,2020年,Seddon等人,2020年)。这种方法可能导致不适应的做法,特别是考虑到迅速转向以生物多样性为基础的恢复力为关键的战略。以碳为中心的方法也不利于现存森林的保护,特别是成熟的原始生长林,并减少了促进森林持久性的努力(Lewis等人,2019年)。
我们强调,次生林是土地管理的一种重要的高成本效益选择,可支持拉丁美洲目前代表性不足的多种环境承诺(Chazdon和Guariguata 2016, 2018)。我们给出了三个理由:
- 次生林中的树木已经在快速生长并吸收和储存碳(Asner et al. 2010, Chazdon et al. 2016, Poorter et al. 2016),而新种植的森林需要很多年才能大量吸收二氧化碳(框1-3;图2 a)。此外,次生林有更大的树种多样性和更高的固碳率,而人工林往往是单一栽培和存储量,固碳量较少,比次生林慢,特别是在亚马逊地区。由于二氧化碳去化率需要迅速提高,才能达到温度限制目标(IPCC 2018),因此次生林是吸收大气碳的快速途径。更频繁的干旱、虫害和疾病会影响种植园和农林业企业的成功建立及其相关效益。
- 人工林和农林复合系统大多是出于商业目的而种植的,而不是为了产生永久的森林覆盖。因此,这些森林可能会损害长期的碳封存,因为树木在几十年后被收割(Farley et al. 2005, Luzar 2007, Raboin and Posner 2012)。事实上,我们没有发现任何报告种植园或农林复合系统超过30年的研究。社会经济和环境方面的冲突也影响到人类管理森林的持久性。例如,在库斯科的一个农村社区(Piñapampa),每周大约有80-90辆卡车送货桉树将木材运到镇上用作燃料(从而向大气中释放二氧化碳),而当地居民在一个典型的伐木季节平均能赚6万到8.1万美元(Luzar 2007)。考虑到成本和效益,这些价值远远高于基于碳封存的潜在市场(Luzar 2007, rabin和Posner 2012)。
- 为了优化造林和再造林的效益,经常需要大量的土地,这可能会破坏自然生境,并与粮食生产、城市空间和其他土地用途产生竞争。例如,秘鲁安第斯山脉的大片天然草原上长满了奇异植物桉树而且松果体对水文服务和生物多样性保护产生负面影响的物种(Krois和Schulte 2013, Tovar等人2013,Bonnesoeur等人2019)。
次生林作为生物多样性保护的生境
次生林不仅是碳的有效和具有成本效益的天然吸收体,而且还为许多动植物,包括人类管理的森林中经常缺少的受威胁物种提供了必要的栖息地。这一功能在高度地方性的生态系统中尤为重要,例如安第斯山脉和沿海的半落叶干燥森林。随着时间的推移,自然再生的森林正在经历自组织、结构复杂性和多样性的增加,而人工林和农林复合系统则经历着持续的人为干预。虽然管理森林比裸地更好,但与次生林相比,管理森林的保护目标可能会打折扣。反过来,生物多样性为人类提供了基本服务,包括降低人畜共患病的风险(Gibb等人,2020年)。秘鲁已经实现了其生物多样性战略计划(2011-2020年)的各种目标,这些目标与《生物多样性公约》下的爱知生物多样性保护目标有关:例如,目标11,其中规定到2020年必须保护至少17%的陆地地区(MINAM 2015年,Maxwell等人2020年)。然而,气候变化的复杂性要求从新的角度beplay竞技看待保护战略,不仅包括永久性保护区,还包括与走廊和临时保护区相结合,以创建灵活的网络,为生物多样性重新分布提供关键过渡区域(D’aloia等,2019年)。在目前具有异质性生境马赛克的景观中,次生林是连接保护区的基础,也可能提供区域生境异质性和植被结构复杂性。在此基础上,Horváth等人(2019)强调,连接丧失导致的栖息地丧失加剧了物种丰富度的损失,从而加强了栖息地连接在生物多样性保护政策中的重要性。次生林的持续存在对生物多样性的长期保护至关重要,而长期保护又有助于保护生物多样性为人类提供的多种服务,首先是预防流行病和人畜共患疾病。
次生林在水文服务中的作用
供水是21世纪社会面临的关键问题之一。由于森林砍伐和气候变化改变了天气模式及其可beplay竞技变性,严重的缺水和洪水可能会变得更加频繁。在这种情况下,植被恢复可以对当地和区域的供水和调节产生积极或消极的影响。就秘鲁而言,次生林影响着当地和区域的供水,这在安第斯山脉尤其重要。安第斯地区不仅是当地社区的主要水源(雨水和冰川融水),也是太平洋沿岸干旱地区(Beresford-Jones 2004, Whaley et al. 2010)和亚马逊东部地区(Clark et al. 2014, Doornbos 2015)的主要水源。因此,生物多样性和数百万人的生计直接依赖安第斯生态系统提供的水服务(Doornbos 2015年)。此外,水对农业、能源、渔业和旅游业等重要经济活动至关重要。在秘鲁,70%的水力发电用水来自安第斯河(Doornbos 2015)。热带安第斯山脉的温度一直在上升,冰川融化,下游供水增加(Vuille等人,2008,2018)。然而,水的增加是暂时的,是不可持续的。 In contrast, future reductions in water availability for both human consumption and other economically important activities is expected (Vuille et al. 2018). The imminent impacts of climate change on water supply motivated the Peruvian government and nongovernmental organizations to implement a variety of rainwater harvesting techniques, including forestation (Somers et al. 2018, Locatelli et al. 2020). Exotic trees (桉树而且松果体)在安第斯山脉的大面积种植对水文服务有负面影响,减少了河流流量和地下水补给,从而减少了下游用户的可用水(Luzar 2007, Ochoa-Tocachi et al. 2016, Ochoa-Tocachi 2019)。此外,快速增长的异国情调桉树而且松果体种植园建立在湿地或安第斯集水区的边缘,导致了上述负面影响(Luzar 2007, Ochoa-Tocachi et al. 2016)。因此,为了实现预期的目标,有必要重新调整国家恢复战略。应避免在天然草地上种植外来树种,提倡原生物种次生林的保持和恢复以及成熟森林的保护,以促进其在安第斯山脉的良好水文控制能力(专栏2)。
尽管我们倾向于强调外来树种对水分产量的负面影响,但如果管理得当(例如,在远离流域的退化土地等适当地区实施),它们可以发挥重要的经济作用,并有助于控制侵蚀和稳定土壤,防止滑坡(Guns和Vanacker 2013年,Bonnesoeur等人2019年)。此外,种植园可以通过避免对原生森林的进一步砍伐,间接增强一些生态系统服务(Bonnesoeur等,2019年)。最后,重要的是要考虑到水文调节的改善是一个缓慢的过程,可能需要几十年的时间,使社会和环境的成果难以实现(Bonnesoeur等,2019年)。
结束语
我们提供了关于森林过渡的生态系统服务和危害的已发表证据的有力综合,以鼓励采取促进秘鲁景观次生林的可持续利用和保护的政策。在这篇综述中,我们收集了经验证据,为未来的研究和政策决策提供了一个框架(图4A)。我们发现,森林转型在抵消碳排放、调节水文服务和为生物多样性提供栖息地方面提供了重要服务。然而,生态系统服务的数量和重要性因森林过渡类型而异。次生林通过维持景观连通性、区域异质性和植被结构复杂性,提供从减缓气候变化和调节水量到物种保护等多种具有成本效益的服务。beplay竞技
我们的研究结果突出了依靠次生林来实施更雄心勃勃的综合环境倡议的机会,如《生物多样性公约-爱知生物多样性目标》、《联合国气候变化框架公约- ndc - redd +》和土地恢复倡议20x20。目前的承诺包括高成本战略,包括潜在的危害(例如,根据20x20倡议,200万商业树木种植园)。相比之下,次生林是一种低成本、短期、高效的战略,可缓解气候和生物多样性危机,并最终为社会提供重要的生态系统服务,例如供水。我们敦促既考虑生态系统的服务,也考虑生态系统的危害,并评估每个特定区域不同土地选择的相对重要性(图4),以避免实现一个目标会损害另一个目标的权衡。政策制定者和实践者可以使用我们的综合方法来支持在秘鲁的地方和区域范围内实现特定最终目标的最佳过渡类型的决策。我们的综合研究有助于提高可见度,并强调保护次生林和成熟挺立林的战略的有用性。最后,还必须考虑到次生林的持续存在与成本和效益密切相关,当地人在保护林区方面的利益可能是决定性的(图4A)。
作者的贡献
RT, NS和RMRC提出了这个想法;RT进行了审查;RT、NS、EGC、TEBE和RMRC撰写了草案;所有作者都对最终版本的手稿进行了讨论和贡献。
致谢
这项工作得到了世界银行赠款(合同011-2019- fondecyte - bm - inco - inv) CONCYTEC(秘鲁)的支持。我们感谢José C. Silva博士和三位匿名审稿人对手稿早期草稿提出的有用建议。
数据可用性
本稿所用的所有数据见附录1和附录2。
文献引用
Aerts, R., S. Spranghers和Ç。h .Şekercioğlu。2017.对生态系统服务的保护并不能保证对秘鲁树荫咖啡景观中的鸟类的保护。国际鸟类保护27(1):71-82。https://doi.org/10.1017/S0959270916000149
艾德,t.m., H. R.格劳,J.格雷瑟,M. J. Andrade-Nuñez, E. Aráoz, A. P.巴罗斯,M.坎波斯-塞奎拉,E.查康-莫雷诺,F.奎斯塔,R.埃斯皮诺萨,M.佩拉尔沃,M. H.波尔克,X.鲁埃达,A.桑切斯,K. R.杨,L. Zarbá, K. S.齐默尔。2019。2001 - 2014年热带和亚热带安第斯地区木本植被动态:卫星图像解译和专家验证全球变化生物学25(6):2112-2126。https://doi.org/10.1111/gcb.14618
阿雷格里,J. L. Arévalo,和A. Ricse. 2003。黑葡萄酒según秘鲁亚马逊地区的铁地和土地。在m.d. Sánchez m.r. Méndez,编辑。Agroforestería para la producción animal en América拉丁- II -纪念la segunda conferencia electrónica (Agosto de 2000-Marzo de 2001)。粮农组织,罗马,意大利。https://www.fao.org/3/Y4435S/Y4435S00.htm
Álvarez-Berríos, N. J. L 'Roe和L. Naughton-Treves。2021.将手工采金正式化能减轻对环境的影响吗?秘鲁亚马逊的森林砍伐证据。环境研究通讯16(6):064052。https://doi.org/10.1088/1748-9326/abede9
Aragón, S, N.萨利纳斯,A.尼娜-奎斯佩,V. H.奎伦,G. R.包卡尔,W.华曼,P. C. Porroa, J. C.奥拉特,R.克鲁兹,J. G. Muñiz, C. S.尤帕伊卡纳,T. E. B.埃斯皮诺萨,R.蒂托,E. G.科西奥,R. M.罗曼-奎斯塔。2021.秘鲁次生山森林的地上生物量:农林业遗产的碳恢复缓慢。全球生态与保护28:e01696。https://doi.org/10.1016/j.gecco.2021.e01696
Arévalo-Gardini, E. M. Canto, J. Alegre, O. Loli, A. Julca,和V. Baligar. 2015。长期土壤理化性质的变化改善了秘鲁亚马逊地区可可基因型的自然和传统农林复合管理制度。《公共科学图书馆•综合》10 (7):e0132147。https://doi.org/10.1371/journal.pone.0132147
阿斯纳,G. P, D. E. Knapp, R. E. Martin, R. Tupayachi, C. B. Anderson, J. Mascaro, F. sina, K. D. Chadwick, S. Sousan, M. Higgins, W. Farfan, M. R. Silman, W. A. Llactayo León, A. F. Neyra Palomino. 2014。La geografía del carbono en alta resolución del Perú。卡内基科学研究所,华盛顿特区,美国。https://geoservidorperu.minam.gob.pe/geoservidor/Archivos/Documentos/CarnegiePeruCarbonReport-Spanish.pdf
阿斯纳,G. P., G. V. N.鲍威尔,J.马斯卡罗,D. E.纳普,J. K.克拉克,J.雅各布森,T.肯尼迪-鲍登,A.巴拉吉,G.佩斯-阿科斯塔,E.维多利亚,L.塞卡达,M.瓦尔基,和R. F.休斯。2010。亚马逊高分辨率森林碳储量和碳排放。国家科学院院刊107(38):16738-16742。https://doi.org/10.1073/pnas.1004875107
巴尔塔扎尔Peña, a.l. 2011。炭烧aéreo almacenado en una plantación de bolaina blanca (Guazuma crinita Martius) de un año de edad, en el centro poblado Nuevo Edén, alto Madre de Dios。论文。国家农业大学,廷戈María, Perú。http://repositorio.unas.edu.pe/handle/UNAS/563
Barbaran, g . 1998。Determinación在佛得角的生态环境中,碳的生物和我们的原则。论文。乌卡亚利国立大学,普卡尔帕,Perú。
Barbier, E. B. J. C. Burgess, A. Grainger. 2010。森林转型:向更全面的理论框架迈进。土地政策27(2):98-107。https://doi.org/10.1016/j.landusepol.2009.02.001
J。巴洛,t·a·加德纳i s Araujo t . c . Avila-Pires a . b . Bonaldo J·e·科斯塔·m·c·埃斯波西托l . v .费雷拉,J·霍斯,m . i m·埃尔南德斯m . s . Hoogmoed r·n·雷特n . f . Lo-Man-Hung J·r·马尔科姆·m·b·马丁斯l . a . m .城区r . Miranda-Santos a . l . Nunes-Gutjahr w . l .扶持l·帕里s . l .彼得斯·m·a . Ribeiro-Junior m·n·f·达席尔瓦c·达席尔瓦莫塔和c·a·佩雷斯。2007。热带原生林、次生林和人工林生物多样性价值的量化。国家科学院学报104(47):18555-18560。https://doi.org/10.1073/pnas.0703333104
Bennett-Curry, A., Y. Malhi和M. Menton, 2013。自然资源供应链中的泄漏效应:来自秘鲁商业木炭市场的案例研究。国际可持续发展与世界生态学报20(4):336-348。https://doi.org/10.1080/13504509.2013.804892
贝雷斯福德-琼斯博士,2004。着Prosopis秘鲁南海岸的人类关系:下伊卡河谷环境和文化轨迹背景下的河岸森林。论文。剑桥大学,英国剑桥。
伯纳切·杰西,2019年。Valoración económica我们的二氧化碳和植物的安全stro de CO2 (蓝桉labil) y pino (松果体放射虫纲) 11 años华卡尔县科查塔马市安博省Huánuco - Setiembre, 2018 - 2019年2月。论文。德大学Huánuco, Huánuco, Perú。http://repositorio.udh.edu.pe/123456789/1776
Mathez-Stiefel。2019.安第斯山脉森林和造林对水文服务的影响:系统综述。森林生态与管理433:569-584。https://doi.org/10.1016/j.foreco.2018.11.033
L.布瑞尔,P.劳,B.德维特,D.拉巴特,W.拉瓦多,A.库托,A.维拉,A.阿尔瓦拉多,J. Ordoñez。2015.秘鲁太平洋沿岸北部至中部ENSO与降水关系的低频调制及其趋势。水文过程29(6):1252 - 1266。https://doi.org/10.1002/hyp.10247
布兰登,k . 2014。热带森林的生态系统服务:当前科学综述。全球发展中心工作文件380。全球发展中心,美国华盛顿特区。https://doi.org/10.2139/ssrn.2622749
Bruijnzeel,洛杉矶,2004年。热带森林的水文功能:看不到树木的土壤?农业生态与环境44(1):1 - 5。https://doi.org/10.1016/j.agee.2004.01.015
布洛克,E. L., C. E.伍德科克,P. Olofsson。利用光谱解混和Landsat时间序列分析监测热带森林退化。环境遥感238:110968。https://doi.org/10.1016/j.rse.2018.11.011
巴特勒,R. A.和W. F.劳伦斯,2009。油棕是亚马逊雨林的下一个威胁吗?热带保护科学2(1):1-10。https://doi.org/10.1177%2F194008290900200102
布达特,W., V. Iñiguez和B.德Bièvre。2007.安第斯山区造林和栽培对水分产量的影响páramo。森林生态与管理51(2):22-30。https://doi.org/10.1016/j.foreco.2007.06.035
Caballero Espejo, J., M. Messinger, F. Román-Dañobeytia, C. Ascorra, L. E. Fernandez, M. Silman. 2018。秘鲁亚马逊地区金矿开采导致的森林砍伐和森林退化:34年展望遥感10(12):1903。https://doi.org/10.3390/rs10121903
考德威尔,M. M.道森,J. H.理查兹,1998。水力提升:水从植物根部流出的结果。环境科学,113(2):151 - 161。https://doi.org/10.1007/s004420050363
M. M.考德威尔和J. H.理查兹。1989。液压提升:上部根系的出水提高了深层根系的吸水效率。环境科学,79(1):1 - 5。https://doi.org/10.1007/BF00378231
Cerrón, J., J. del Castillo, S. L. Mathez-Stiefel,和E. Thomas. 2017。经验的Lecciones aprendidas de experience de restauración en el Perú。生物多样性国际组织,秘鲁利马。https://www.bioversityinternational.org/e-library/publications/detail/lecciones-aprendidas-de-experiencias-de-restauracion-en-el-peru/
Cerrón, J., T. Fremout, R. Atkinson, E. Thomas和J. Cornelius. 2019。我们的经验restauración我们的国家,我们的热带国家,我们的北方,我们的国家Perú,我们的国家,我们的国家。生物多样性国际组织,秘鲁利马。https://www.bioversityinternational.org/e-library/publications/detail/experiencias-de-restauracion-y-fuentes-semilleras-en-el-bosque-seco-tropical-del-norte-del-peru-est/
Chausson, A., B. Turner, D. Seddon, N. Chabaneix, C. A. J. Girardin, V. Kapos, I. Key, D. Roe, A. Smith, S. Woroniecki和N. Seddon. 2020。绘制基于自然的气候变化适应解决方案的有效性图。beplay竞技全球变化生物学26(11):6134-6155。https://doi.org/10.1111/gcb.15310
Chávez苏亚佐,J. P. 2018。捕获北方海岸(Lambayeque),科莫mitigación前线,坎比奥climático。论文。秘鲁利马的国立农业大学(Universidad national Agraria la Molina)。https://hdl.handle.net/20.500.12996/3346
Chavez-Villavicencio, c . 2013。la aves de la ciudad de Piura, Perú y sus alrededores: ocho años mirando al cielo。生物学家11(2):193 - 204。https://doi.org/10.24039/rtb2013112398
Chazdon, r·L。e . n . Broadbent d·m·a . Rozendaal f . Bongers a·m·a·摩尔诺t . m .助手,p . Balvanera j . m . Becknell诉Boukili p h . s . Brancalion d·克雷文j . s . Almeida-Cortez g·a·L·卡布拉尔德容,j . s . Denslow d·h·削弱s . j . DeWalt j . m . Dupuy称:"现在s·m·杜兰·m·m·圣埃斯皮里图m . c . Fandino r·g·塞萨尔j·s·霍尔,j·L·Hernandez-Stefanoni c . c . Jakovac a b,琼奇拉d . Kennard s . g .莱彻m . Lohbeck m . Martinez-Ramos p . Massoca j . a . Meaver . Mesquita f•莫拉r·穆尼奥斯r . Muscarella y . r . f . Nunes s Ochoa-Gaona e . Orihuela-Belmonte m . Pena-Claros e·a·Perez-Garcia d . Piotto j . s .权力,j . Rodriguez-Velazquez即Romero-Perez, j . Ruiz j·g . Saldarriaga sanchez - azofeifa告诉本网络记者,n . b . Schwartz m·k。施n g·斯文森,m . Uriarte m . van Breugel h . van der细胞膜,m·d·m·维罗索h .背心i c·g·维埃拉t . v .便当,g·b·威廉姆森,2016 l .要隘。。拉丁美洲热带次生林再生的固碳潜力科学进步2 (5):e1501639。https://doi.org/10.1126/sciadv.1501639
查兹登,R. L.和M. R.瓜里瓜塔,2016。自然更新作为热带地区大规模森林恢复的工具:前景与挑战。时48(6):716 - 730。https://doi.org/10.1111/btp.12381
查兹登,R. L.和M. R.瓜里瓜塔。森林景观恢复决策支持工具的现状与展望。偶尔纸183。国际林业研究中心,茂物,印度尼西亚。https://doi.org/10.17528/cifor/006792
查兹登,R. L., D. Lindenmayer, M. R. Guariguata, R. Crouzeilles, J. M. Rey Benayas和E. Lazos Chavero. 2020。通过经济和政策干预,在以前的农业用地上促进天然林的更新。环境研究通讯15(4):043002。https://doi.org/10.1088/1748-9326/ab79e6
丘奎鲁塔,p.d., G. F.鲁伊斯,C. T.萨拉斯,l.a. López。2016.炭黑的almacenado,在我们的地岛上,在región San Martín Perú。Rinderesu 1(2): 57 - 67。http://rinderesu.com/index.php/rinderesu/article/view/22
克拉克,K. E, M. A.托雷斯,A. J.韦斯特,R. G.希尔顿,M.纽,A. B.霍沃斯,J. B.费雪,J. M.拉普,A.罗伯斯·卡塞雷斯,Y.马尔希。2014。热带安第斯森林集水区的水文状况。水文与地球系统科学18(12):5377-5397。https://doi.org/10.5194/hess-18-5377-2014
气候行动追踪,2019年。秘鲁国家决定贡献(NDC)评估。气候行动网络,波恩,德国。https://climateactiontracker.org/countries/peru/2019-09-19/
科罗拉多·祖卢阿加,G. J.和A. D.罗德瓦尔德,2015。安第斯山区混种鸟群对生境变化和森林砍伐的响应生物保护188:72 - 81。https://doi.org/10.1016/j.biocon.2015.02.008
康夏,J. Y.阿雷格里,J. C.阿雷格里,V.波科莫查。2007。Determinación森林生态系统aérea森林系统Theobroma可可L. en el departamento de San Martin, Perú。Ecologia Aplicada 6(1 - 2): 75 - 82。http://www.scielo.org.pe/scielo.php?pid=S1726-22162007000100009&script=sci_abstract
Csillik, O.和G. P. Asner. 2020。秘鲁亚马逊地区金矿开采产生的地上碳排放。环境研究通讯15(1):014006。https://doi.org/10.1088/1748-9326/ab639c
奎利亚尔·鲍蒂斯塔,J. E.和E. J.萨拉查·伊诺斯托萨。2016。Dinámica在不同的体系里,有不同的体系,有不同的体系,有不同的体系,有不同的体系,有不同的体系,有不同的体系,有不同的体系mitigación,有不同的体系,有不同的体系,mitigación。国家研究所Innovación阿格拉里亚,利马,秘鲁。http://repositorio.inia.gob.pe/handle/20.500.12955/363
L.克勒特,D. J.曼,F. J. J. Muñoz海曼(E. W. Heymann)。狨猴和蜣螂:秘鲁亚马逊地区一种高效的双杂食传播系统。时43(1):84 - 92。https://doi.org/10.1111/j.1744-7429.2010.00655.x
D 'Almeida, C. J. Vörösmarty, G. C. Hurtt, J. A. Marengo, S. L. Dingman和B. D. Keim. 2007。亚马逊森林砍伐对水文循环的影响:规模和分辨率的研究进展。气候科学进展27(5):633-647。https://doi.org/10.1002/joc.1475
达洛亚、c.c.、I.诺约凯蒂斯-刘易斯、C.布莱克福德、C.朱、J. M. R.柯蒂斯、E.达林、F.吉查、S. J.勒鲁、A. C.马腾森、B.雷菲尔德、J. M. Sunday、A.雪瑞布和M. j。福丁》2019。气候变化下生物多样性保护的永久性保护区与动态保护区耦合网络beplay竞技生态学与进化前沿7:27。https://doi.org/10.3389/fevo.2019.00027
de Valença, A. W. S. J. Vanek, K. Meza, R. Ccanto, E. Olivera, M. Scurrah, E. A. Lantinga,和S. J. Fonte. 2017。土地利用是秘鲁安第斯山脉中部农业景观土壤肥力和生物多样性的驱动因素。生态学应用27(4):1138 - 1154。https://doi.org/10.1002/eap.1508
德尔加多·帕雷德斯,G. E. C. Vásquez-Díaz, F. Tesén-Núñez, B. Esquerre-Ibañez, F. Z. Da-Silva,和C.罗哈斯-伊德罗戈。2020.Vegetación arbórea del Cerro Tres Puntas de Pilasca,(萨拉斯-莫图佩),兰巴耶克,Perú。墨西哥林业科学通报11(58):154-181。https://doi.org/10.29298/rmcf.v11i58.549
差一点,b . 2015。El valor de los bosques andinos en asegurar agua y suelo en contesto de creciente riesgo climático:¿(re)conocemos lo不可听?第三条意见。Bosques Andinos项目,秘鲁利马。https://www.bosquesandinos.org/wp-content/uploads/2015/12/061115_articulo_n3.pdf
Duran-Bautista, E. H, I. Armbrecht, A. N. Serrão acoli, J. C. Suárez, M. Romero, M. Quintero和P. Lavelle. 2020。白蚁对改良亚马逊景观土壤生态系统服务功能的指示作用117:106550生态指标。https://doi.org/10.1016/j.ecolind.2020.106550
Ehrenbergerová, L., E. Cienciala, A. kuevera, L. Guy和H. Habrová。2016.秘鲁维拉黎加不同荫凉树木的农林咖啡种植园的碳储量。农林复合经营系统90(3):433 - 445。https://doi.org/10.1007/s10457-015-9865-z
埃里森、M. N.富特和K.毕晓普。2012。关于森林覆盖-水产量的辩论:从需求到供给侧的思考。全球变化生物学18(3):806-820。https://doi.org/10.1111/j.1365-2486.2011.02589.x
埃斯科巴尔,J.和M.托雷罗,2003。秘鲁不利的地理位置和不同的福利。77 - 121页在R.坎伯尔和A. J.维纳布尔斯,编辑。空间不平等与发展。牛津大学出版社,英国牛津。https://doi.org/10.1093/0199278636.003.0005
Escobedo Grandez, A. 2021。agonegocios y危机climática en el Perú:单眼的,deforestación y碳碳的排放物amazonía秘鲁。乐施会América,秘鲁利马。https://peru.oxfam.org/latest/policy-paper/agronegocios-y-crisis-climatica-en-peru
法利,E. G. Jobbágy, R. B.杰克逊,2005。造林对产量的影响:对政策有影响的全球综合。全球变化生物学11(10):1565-1576。https://doi.org/10.1111/j.1365-2486.2005.01011.x
法利,K. A., E. F.凯利,R. G. M.霍夫斯泰德,2004。厄瓜多尔安第斯山脉草原转变为松树种植园后的土壤有机碳和水分保持。生态系统7(7):729 - 739。https://doi.org/10.1007/s10021-004-0047-5
Fitts, l.a, Z. A. Cruz-Burga和M. La Torre-Cuadros。2020.秘鲁亚马逊地区的野生橡胶开采:作为决策工具的当地认知和社会经济指标。人种生物学与保护9:24。https://doi.org/10.15451/ec2020-06-9.24-1-26
联合国粮食及农业组织。2016.博斯奎斯和埃尔坎比奥climático:特拉巴霍文件14。粮农组织,罗马,意大利。https://www.fao.org/documents/card/en/c/50d7476c-8ac1-40d8-a704-db7ed0281662/
联合国粮食及农业组织。2020.2020年全球森林资源评估:主要报告。粮农组织,罗马,意大利。https://www.fao.org/documents/card/en/c/ca9825en
加利斯,G. R., G. Larrea-Gallegos和I. Vázquez-Rowe。2019.秘鲁亚马逊地区公路扩建的威胁。羚羊53(2):284 - 292。https://doi.org/10.1017/S0030605317000412
加玛拉·拉莫斯,2001。Estimación del contenido de carbono en plantaciones de蓝桉Labill, en Junin, Perú。在国际Simposio medición森林生态系统碳碳监测。墨西哥奇瓦瓦,奇瓦瓦大学Autónoma。
加西亚、c.a、S. Savilaakso、R. W. Verburg、V. Gutierrez、S. J.威尔逊、C. B.克鲁格、M.萨森、B. E.罗宾逊、H.莫尔斯伯格、B.纳伊米、J. M. Rhemtulla、H. Dessard、V. Gond、C. Vermeulen、F. Trolliet、J. Oszwald、F. Quétier、S. A. Pietsch、J. f。巴斯廷,A.德雷,M. B. Araújo, J.加扎乌尔,P. O.韦伯,2020。全球森林转型是人类的事情。一个地球2(5):417 - 428。https://doi.org/10.1016/j.oneear.2020.05.002
加文,2004年。秘鲁亚马逊森林利用价值的生态演替变化及其对土地管理的意义保护生物学18(6):1562 - 1570。https://doi.org/10.1111/j.1523-1739.2004.00241.x
加文,2007年。休耕地的觅食:秘鲁亚马逊连续连续的狩猎模式。生物保护134(1):64 - 72。https://doi.org/10.1016/j.biocon.2006.07.011
Giam x 2017。热带森林砍伐导致全球生物多样性丧失。中国科学院学报114(23):5775-5777。https://doi.org/10.1073/pnas.1706264114
吉布,R., D. W.雷丁,K. Q. Chin, C. A.唐纳利,T. M.布莱克本,T.纽博尔德,K. E.琼斯,2020年。在人类主导的生态系统中,人畜共患病宿主多样性增加。自然584(7821):398 - 402。https://doi.org/10.1038/s41586-020-2562-8
“德尔秘鲁。2016.contribución国家大会Perú -国家大会:负责任的议程climáticamente。秘鲁政府,秘鲁利马。https://www.minam.gob.pe/cambioclimatico/wp-content/uploads/sites/11/2015/12/LA-CONTRIBUCI%c3%93N-NACIONAL-DEL-PER%c3%9a1.pdf
“德尔秘鲁。2020.国家决定性贡献Perú: 2021-2030年报告actualización。秘鲁政府,秘鲁利马。https://cdn.www.gob.pe/uploads/document/file/1675213/Reporte%20de%20
Actualizaci % C3%B3n % 20 de % 20 las % 20 ndc % 20 del % 20 / % C3%BA % 20 al % 202030. pdf
冈萨雷斯,P., B. Kroll, C. R. Vargas. 2014。秘鲁中部塞尔瓦的热带雨林生物多样性和地上碳变化与不确定性。森林生态与管理312:78-91。https://doi.org/10.1016/j.foreco.2013.10.019
Gonzales-Zuniga, s . 2018。国家决定捐款的执行情况:秘鲁国家报告。德国Umweltbundesamt, Dessau-Roß刘。https://www.umweltbundesamt.de/sites/default/files/medien/1410/publikationen/2018-11-30_climate-change_31-2018_country-report-peru.pdf
《格里斯科姆》,B. W.、J.亚当斯、P. W.埃利斯、R. A.霍顿、G.洛马克斯、D. A.米特娃、W. H.施莱辛格、D.索奇、J. V. Siikamäki, P.史密斯、P.伍德伯里、C. Zganjar、A.布莱克曼、J.金帕里、R. T.科南特、C.德尔加多、P.埃利亚斯、T.戈帕拉克克里希纳、M. R.哈姆西克、M.埃雷罗、J.基塞克、E.兰迪斯、L.莱维特、S.明尼迈耶、S.波拉斯基、P.波塔波夫、F. E.普茨、J.桑德曼、M.西尔维乌斯、E.渥伦伯格、J.法乔内。2017。自然气候解决方案。中国科学院学报114(44):11645-11650。https://doi.org/10.1073/pnas.1710465114
《枪炮》,M.和V. Vanacker, 2013。热带安第斯山脉森林覆盖变化轨迹及其对滑坡发生的影响环境科学与技术,29(6):359 - 359。https://doi.org/10.1007/s12665-013-2352-9
Gurmendi Porras, C. G.和W. B. Orihuela Izaguirre. 2019。Valoración económica de la reserva de carbono deEscallonia resinosa(Ruiz &奶油水果蛋白饼。)珀耳斯。en圣佩德罗德Saños y Pucará。秘鲁万卡约国家中心大学Perú。http://hdl.handle.net/20.500.12894/5320
汉森,m.c., P. V. Potapov, R. Moore, M. Hancher, S. A. Turubanova, A. Tyukavina, D. Thau, S. V. Stehman, S. J. Goetz, T. R. Loveland, A. Kommareddy, A. Egorov, L. Chini, c.o. Justice和J. R. G. Townshend. 2013。21世纪森林覆盖变化的高分辨率全球地图。科学》342(6160):850 - 853。https://doi.org/10.1126/science.1244693
哈登,c.p, J.哈特西格,K. A.法利,J.李,L. L.布雷默。2013。土地利用变化对安第斯páramo草地土壤水分的影响。美国地理学家协会年鉴103(2):375-384。https://doi.org/10.1080/00045608.2013.754655
R. Heilmayr, C. Echeverría和E. F. Lambin. 2020。智利森林补贴对森林覆盖、碳和生物多样性的影响。自然的可持续性3(9):701 - 709。https://doi.org/10.1038/s41893-020-0547-0
Holl, K. D.和P. H. S. Brancalion. 2020。植树不是一个简单的解决方案。科学》368(6491):580 - 581。https://doi.org/10.1126/science.aba8232
Hölscher, D. J.麦康森和J.- m。罗伯茨。2005。潮湿热带地区的森林恢复:植被结构、营养池和水文循环的变化。598 - 621页在M.博内尔和L. A. Bruijnzeel,编辑。潮湿热带地区的森林、水和人:土地和水综合管理的过去、现在和未来水文研究。剑桥大学出版社,英国剑桥。https://doi.org/10.1017/CBO9780511535666.031
霍根,2009。入侵和撤退:在秘鲁中部不断变化的农业景观中蜣螂的移动组合。生物多样性与保护18(13):3519。https://doi.org/10.1007/s10531-009-9658-7
Horváth, Z., R. Ptacnik, C. F. Vad和J. M. Chase. 2019。60多年来,由于景观连接性的变化,栖息地的丧失加速了区域和当地生物多样性的丧失。生态学通讯22(6):1019 - 1027。https://doi.org/10.1111/ele.13260
霍斯纳,P. A., M. J.安德森,M. B.罗宾斯,A. urbayo - tello, L. Cueto-Aparicio, K. Verde-Guerra, L. Sánchez-González, A. G. Navarro-Sigüenza, R. L.博伊德,J. Núñez, J. Tiravanti, M. Combe, H. L.欧文斯,A. Townsend Peterson. 2015。秘鲁阿亚库乔省和库斯科省Apurímac河上游的鸟类调查:新的分布记录和生物地理学、分类学和保护意义。鸟类学报127(4):563-581。https://doi.org/10.1676/14-178.1
国际农林研究中心。1998.对新需求的回应tecnológicas,对农业工业的新需求investigación。1996年末报告,1998年初三报告。德萨罗洛美洲银行和国际农林研究中心,秘鲁利马。
政府间气候变化专门委员会。beplay竞技2018.为决策者Sumary。页面3-24在V.马森-德尔莫特、P.翟鹏、H. O. Pörtner, D.罗伯茨、J.斯卡、P. R.舒克拉、A.皮拉尼、W. Moufouma-Okia、C. Péan, R.皮德考克、S.康纳斯、J. B. R.马修斯、陈扬、周x、M. I.戈米斯、E.朗诺伊、T.梅科克、M.提诺和T.沃特菲尔德编辑。全球变暖1.5摄氏度。在加强全球应对气候变化威胁、可持续发展和消除贫困的努力的背景下,政府间气候变化专门委员会关于全球变暖比工业化前水平高1.5°C的影响和相关全球温室气体排放途径的特别报告。beplay竞技剑桥大学出版社,英国剑桥。https://doi.org/10.1017/9781009157940.001
Jezeer, R. E., M. J. Santos, P. A. Verweij, R. G. A. Boot和Y. Clough, 2019。秘鲁咖啡农林复合系统在不降低产量的情况下对多种生态系统服务的好处。生态系统服务40:101033。https://doi.org/10.1016/j.ecoser.2019.101033
琼斯,i.l., S. J.德沃尔特,O. R.洛佩兹,L.邦内菲尔德,Z.帕蒂森和D. H.登特,2019。次生林地上、地下碳储量呈解耦关系,与林龄和土壤养分呈正相关。总环境科学697:133987。https://doi.org/10.1016/j.scitotenv.2019.133987
克罗斯,J.和A.舒尔特,2013。模拟秘鲁安第斯山脉北部朗基洛流域水土保持措施的水文响应。147 - 184页在水与环境动力学:第六届水资源与环境研究国际会议:论文集。世界气象组织,德国科布伦茨。https://doi.bafg.de/BfG/2014/ICWRER_2013.pdf
拉普爱尔、阿雷格里、L. Arévalo。2004.Determinación生物的碳和生物的珍藏aérea,在不同的体系中在San的土地上Martín, Perú。Ecologia Aplicada 3(1 - 2): 35-44。https://doi.org/10.21704/rea.v3i1-2.269
León Cáceres, F. J. 2019。环境的影响,deforestación,我知道”El Algarrobal”de Conache -Laredo, 2016。论文。特鲁希略国立大学,特鲁希略,秘鲁。http://dspace.unitru.edu.pe/handle/UNITRU/12919
刘易斯,S. L, C. E.惠勒,E. T. A.米查德和A.科赫,2019。恢复天然林是去除大气中碳的最好方法。自然568(7750):25 - 28。https://doi.org/10.1038/d41586-019-01026-8
B,右路放倒,人类。Homberger, B. F. Ochoa-Tocachi, V. Bonnesoeur, F. Román, F. Drenkhan和W. Buytaert. 2020。墨西哥的冲击infiltración en el agua y los suelos:¿Qué sabemos?森林趋势,利马,秘鲁。https://www.forest-trends.org/wp-content/uploads/2020/05/Impactos-de-las-zanjas-de-infiltraci%C3%B3n-en-el-agua-y-los-suelos.pdf
Lojka, B., J. Krausová, Š。Kubík, Z. Polesný。2011.秘鲁亚马逊不同土地利用系统昆虫生物多样性评估。103 - 121页在罗哈斯和普列托,编辑。亚马逊盆地:植物、野生动物和环境。新星科学出版社,纽约,纽约,美国。
Luzar, j . 2007。政治生态”森林过渡”:秘鲁安第斯山脉南部的桉树林。民族植物学研究与应用(5):85-93。https://doi.org/10.17348/era.5.0.85-93
麦克唐纳,H.和D.麦肯尼。2020.设想全球森林转型:地位、作用和影响。土地使用政策99:104808。https://doi.org/10.1016/j.landusepol.2020.104808
K.马哈茂德,J.莫里斯,J.科洛比和P.斯拉维奇。2001。巴基斯坦旁遮普省盐碱地农业种植园的地下水吸收和可持续性。农业用水管理48(1):1-20。https://doi.org/10.1016/s0378 - 3774 (00) 00114 - 1
Málaga, N., K. herergoualc 'h, G. Kapp,和C. Martius. 2021。秘鲁亚马逊雨林由受干扰森林转为油棕种植园引起的植被和生态系统碳储量的变化。生态系统24(2):351 - 369。https://doi.org/10.1007/s10021-020-00521-8
Malhi, Y., J. Franklin, N. Seddon, M. Solan, M. G. Turner, C. B. Field和N. Knowlton. 2020。beplay竞技气候变化与生态系统:威胁、机遇和解决办法。英国皇家学会哲学学报B 375:20190104。https://doi.org/10.1098/rstb.2019.0104
Manoli, G., A. Meijide, N. Huth, A. Knohl, Y. Kosugi, P. Burlando, J. Ghazoul, S. Fatichi. 2018。热带森林转为油棕后的生态水文变化。环境研究通讯13(6):064035。https://doi.org/10.1088/1748-9326/aac54e
马伦戈,j.a . 2006。关于亚马逊流域的水文循环:历史回顾和现状。气象学报21(3):1-19。
Martel, C.和L. Cairampoma. 2012。Cuantificación蛋卷蔬菜amazónicas”cicra”, Madre de Dios (Perú)。Ecologia Aplicada 11(1 - 2): 59 - 65。https://doi.org/10.21704/rea.v11i1-2.426
马克斯韦尔,s.l., V.卡扎利斯,N.达德利,M.霍夫曼,A. S. L.罗德里格斯,S.斯托顿,P.维斯康蒂,S.伍德利,N.金斯顿,E.刘易斯,M.马龙,B. B. N.斯特拉斯堡,A.温格,H. D.乔纳斯,O.文特尔,J. E. M.沃森。21世纪的区域保护。自然586(7828):217 - 227。https://doi.org/10.1038/s41586-020-2773-z
麦基,K. M.波特,M.赖特和M.哈吉巴巴埃。利用DNA转移系统分析热带次生林热带土壤无脊椎动物群落组成和丰富度的驱动因素。10:18429科学报告。https://doi.org/10.1038/s41598-020-75452-4
环境部。2014.国家发展战略biológica 2021年:计划acción 2014-2018年。环境部,利马,秘鲁。https://sinia.minam.gob.pe/documentos/estrategia-nacional-diversidad-biologica-2021-plan-accion-2014-2018
环境部。2015.植物麻柏:记忆描述。环境部,利马,秘鲁。https://repositoriodigital.minam.gob.pe/handle/123456789/178
环境部。2016.El Perú y El cambio climático: tercera comunicación national del Perú a la Convención Marco de las nacones Unidas soidas El cambio Climático。环境部,利马,秘鲁。https://sinia.minam.gob.pe/documentos/tercera-comunicacion-nacional-peru-convencion-marco-las-naciones
Moomaw, W. R., S. A. Masino和E. K. Faison. 2019。美国完整的森林:植树造林减缓了气候变化,为最大的利益服务。beplay竞技森林前沿与全球变化2:27。https://doi.org/10.3389/ffgc.2019.00027
迈尔斯,R. A. Mittermeier, C. G. Mittermeier, G. A. B. da Fonseca, J. Kent, 2000。生物多样性热点是保护重点。自然403(6772):853 - 858。https://doi.org/10.1038/35002501
Ochoa-Tocachi, b.f 2019。参与水文监测以支持可持续水资源管理。论文。帝国理工学院,伦敦,英国。https://doi.org/10.25560/75557
奥乔亚-托卡奇,B. F, W.布泰特,B.德Bièvre, R. Célleri, P.克雷斯波,M. Villacís, C. A.勒莱雷纳,L.阿科斯塔,M. Villazón, M.瓜尔帕,J. Gil-Ríos, P.富恩特斯,D.奥拉亚,P. Viñas, G.罗哈斯,S.阿里亚斯。2016。土地利用对热带安第斯流域水文响应的影响。水文过程30(22):4074 - 4089。https://doi.org/10.1002/hyp.10980
Oliveras, I., Y. Malhi, N. Salinas, V. Huaman, E. Urquiaga-Flores, J. Kala-Mamani, J. A. Quintano-Loaiza, I. Cuba-Torres, N. Lizarraga-Morales, r . m。Roman-Cuesta。2014.安第斯山林线附近热带山地云林火灾后森林结构和组成的变化。植物生态多样性7(1-2):329-340。https://doi.org/10.1080/17550874.2013.816800
奥利维拉斯,I., R. M. Román-Cuesta, E.厄奎加-弗洛雷斯,J. A. Quintano Loayza, J.卡拉,V. Huamán, N. Lizárraga, G. Sans, K. Quispe, E. Lopez, D. Lopez, I. Cuba Torres, B. J. Enquist,和Y. Malhi. 2018。热带山地云林的火效应与生态恢复路径全球变化生物学24(2):758-772。https://doi.org/10.1111/gcb.13951
Pécastaing, N.和C. Chávez。2020.的影响厄尔尼诺现象秘鲁北部海岸干旱森林依赖社区福利的现象。生态经济学178:106820。https://doi.org/10.1016/j.ecolecon.2020.106820
佩里,J., B. Lojka, L. G. Quinones Ruiz, P. Van Damme, J. houikka, E. Fernandez Cusimamani. 2016。天然林转换如何影响秘鲁亚马逊地区的昆虫生物多样性:农林复合林业能有所帮助吗?森林7(4):82。https://doi.org/10.3390/f7040082
要隘,L f . Bongers t . m .助手,a . m . Almeyda摩尔诺,p . Balvanera j . m . Becknell诉Boukili p h . s . Brancalion e . n . Broadbent r . L . Chazdon d·克雷文j . s . de Almeida-Cortez g·a·L·卡布拉尔b·h·j·德容j . s . Denslow d·h·削弱s . j . DeWalt j . m . Dupuy称:"现在s·m·杜兰·m·m·圣埃斯皮里图m . c . Fandino r·g·塞萨尔j·s·霍尔,j·L·Hernandez-Stefanoni c . c . Jakovac a b,琼奇拉d . Kennard s . g .莱彻J.-C。Licona, m . Lohbeck e . Marin-Spiotta m . Martinez-Ramos p . Massoca j . a . Meave r . Mesquita f•莫拉r·穆尼奥斯r . Muscarella y . r . f . Nunes s Ochoa-Gaona a . a . de Oliveira e . Orihuela-Belmonte m . Pena-Claros e·a·Perez-Garcia d . Piotto j . s .权力,j . Rodriguez-Velazquez即Romero-Perez, j . Ruiz j·g . Saldarriaga sanchez - azofeifa告诉本网络记者,n . b . Schwartz m·k。施n g·斯文森,m·托莱多m . Uriarte m . van Breugel h . van der细胞膜,m·d·m·维罗索h·f·m·背心A. Vicentini, i.c.g.维埃拉,T. V. Bentos, G. B. Williamson, D. M. A. Rozendaal. 2016。新热带次生林生物量恢复力。自然530(7589):211 - 214。https://doi.org/10.1038/nature16512
M.金特罗,S.文德尔和R. D.埃斯特拉达。2009。提供的服务?模拟安第斯地区的水文和生计,为环境服务计划支付费用。森林生态与管理258(9):1871-1880。https://doi.org/10.1016/j.foreco.2009.04.032
拉宾,M. L.和J. L.波斯纳。2012。松或牧场吗?秘鲁安第斯山脉土地利用变化的估算成本和收益。山地研究与开发32(2):158-168。https://doi.org/10.1659/MRD-JOURNAL-D-10-00099.1
Rasal Sánchez, M. J. Troncos Castro, C. Lizano Durán, O. Parihuamán Granda, D. Quevedo Calle, C. Rojas Idrogo, G. E. Delgado Paredes. 2011。Características edáficas y composición florística La mena y Timbes的墨西哥政府,región Piura, Perú。Ecologia Aplicada 10(1 - 2): 61。https://doi.org/10.21704/rea.v10i1-2.414
Román, F., A. Mamani, A. D. Cruz, C. Sandoval和F. Cuesta. 2018。森林生态系统restauración森林生态系统和森林生态系统vegetación西尔维斯特。sericio国家森林,利马,秘鲁。http://repositorio.serfor.gob.pe/handle/SERFOR/524
Román-Cuesta, R. M. N. Salinas, H. Asbjornsen, I. Oliveras, V. Huaman, Y. Gutiérrez, L. Puelles, J. Kala, D. Yabar, M. Rojas, R. Astete, D. Y. Jordán, M. Silman, R. Mosandl, M. Weber, B. Stimm, S. Günter, T. Knoke,和Y. Malhi. 2011。安第斯云山森林火灾对碳收支的影响:泥炭土壤和树木发芽的重要性。森林生态与管理261(11):1987-1997。https://doi.org/10.1016/j.foreco.2011.02.025
鲁伊斯·罗梅罗,2015年。Ensamblaje y dieta de murciélagos (Chiroptera) en la zona norcidental del Área de Conservación博斯克地区Salitral - Huarmaca, Piura - Perú。论文国立皮乌拉大学,皮乌拉,秘鲁。http://repositorio.unp.edu.pe/handle/UNP/272
萨奇,S. S., N. L.哈里斯,S.布朗,M.莱夫斯基,E. T. A.米查德,W.萨拉斯,B. R.祖塔,W.布尔曼,S. L.刘易斯,S.哈根,S.佩特洛娃,L.怀特,M.西尔曼,A.莫雷尔。2011。三大洲热带地区森林碳储量基准图。中国科学院学报108(24):9899-9904。https://doi.org/10.1073/pnas.1019576108
Saatchi, s.s., R. A. Houghton, R. C. Dos Santos Alvalá, J. V. Soares, Yu. 2007。亚马逊盆地地上活生物量分布。全球变化生物学13(4):816-837。https://doi.org/10.1111/j.1365-2486.2007.01323.x
Scott, d.f, l.a. Bruijnzeel和J. Mackensen, 2005。热带地区造林对水文和土壤的影响。622 - 651页在M.博内尔和L. A. Bruijnzeel,编辑。潮湿热带地区的森林、水和人:土地和水综合管理的过去、现在和未来水文研究。剑桥大学出版社,英国剑桥。https://doi.org/10.1017/CBO9780511535666.032
塞登,N, A. Chausson, P. Berry, C. A. J. Girardin, A. Smith和B. Turner, 2020。理解基于自然的解决方案对气候变化和其他全球挑战的价值和限制。beplay竞技英国皇家学会哲学学报B 375:20190120。https://doi.org/10.1098/rstb.2019.0120
西尔维斯特国家森林动物服务局。2018.经验的de restauración en el Perú: lecciones aprendidas。SERFOR,秘鲁利马。https://www.serfor.gob.pe/portal/wp-content/uploads/2019/01/Experiencias-de-Restauraci%C3%B3n-en-el-Per%C3%BA-Lecciones-aprendidas.pdf
西尔维斯特国家森林动物服务局。2019.通知西尔维斯特国家森林动物名录Perú。SERFOR,秘鲁利马。https://www.serfor.gob.pe/portal/wp-content/uploads/2020/03/INFORME-DEL-INFFS-PANEL-1.pdf
舒克拉、P. R.、J.斯卡、R.斯莱德、R.范·迪门、E.豪伊、J.马利、M.帕塔克和J. P.葡萄牙·佩雷拉主编。2019.技术总结。35 - 74页在舒克拉,斯卡,卡尔沃·布恩蒂亚,马松-德尔莫特,h - o。Pörtner, D. C.罗伯茨、P. Zhai、R. Slade、S. Connors、R. van Diemen、M. Ferrat、E. Haughey、S. Luz、S. Neogi、M. Pathak、J. Petzold、J. Portugal Pereira、P. Vyas、E. Huntley、K. Kissick、M. Belkacemi和J. Malley编辑。beplay竞技气候变化与土地:政府间气候变化专门委员会关于气候变化、荒漠化、土地退化、可持续土地管理、粮食安全和陆地生态系统中的温室气体流动的特别报告。政府间气候变化专门委员会,瑞士日内瓦。beplay竞技https://www.ipcc.ch/site/assets/uploads/sites/4/2020/07/03_Technical-Summary-TS_V2.pdf
史密斯,C. C., F. D. B. Espírito-Santo, J. R.希利,P. J.杨,G. D.伦诺克斯,J.费雷拉和J.巴洛,2020。次生林抵消了巴西亚马逊地区因砍伐森林而产生的碳排放的不到10%。全球变化生物学26(12):7006-7020。https://doi.org/10.1111/gcb.15352
史密斯,C. C., J. R. Healey, E. Berenguer, P. J. Young, B. Taylor, F. Elias, F. Espírito-Santo, J. Barlow. 2021。亚马逊国家的原生林损失和次生林恢复。环境研究通讯16(8):085009。https://doi.org/10.1088/1748-9326/ac1701
Socolar, J. B., E. H. Valderrama Sandoval和D. S. Wilcove. 2019。热带小农农业中被忽视的生物多样性损失。保护生物学33(6):1338 - 1349。https://doi.org/10.1111/cobi.13344
萨默斯,l.d., J. M.麦肯齐,S. C.拉链,B. G.马克,P.拉各斯,M.巴拉尔。2018。在秘鲁安第斯山脉,坡面挖沟是否增加了地下水的补给和基流?水文过程32(3):318 - 331。https://doi.org/10.1002/hyp.11423
索萨·卡斯蒂略,2016年。Valoración económica del secuestro de CO2en tres tipos de boque en el distrito del Alto Nanay, Loreto-Perú-2014。论文。国立大学Amazonía秘鲁,伊基托斯。http://repositorio.unapiquitos.edu.pe/handle/20.500.12737/4283
索托-纳瓦罗,C.、C.拉维尼乌斯,A.阿内尔,X.德拉莫,M.哈富特,S. L.希尔,O. R.沃恩,M.桑托罗,A.布维,S.默莫兹,T.勒托安,夏J., S.刘,元W., S. A.弗里斯,H. K.吉布斯,S.费里尔,T.哈伍德,R.阿尔克梅德,A. M.席珀,G.施密特-特劳布,B.斯特拉斯堡,L.迈尔斯,N. D.伯吉斯,V.卡波斯。绘制碳储存和生物多样性的共同利益,为保护政策和行动提供信息。皇家学会哲学学报b375(1794):20190128。https://doi.org/10.1098/rstb.2019.0128
Srinivas, A.和L. P. Koh. 2016。油棕的扩张导致秘鲁亚马逊Pucallpa地区鸟类数量的下降。全球生态与保护7:183-200。https://doi.org/10.1016/j.gecco.2016.06.005
《时代》,A., M.加里多,E.阿塞韦多,2018。水分关系与生长对干旱胁迫的响应Prosopis tamarugo菲尔。复习一下。土壤与植物营养学报18(2):329-343。https://doi.org/10.4067/S0718-95162018005001103
Timoteo, K, J. Remuzgo, L. Valdivia, F. Sales-Dávila, D. García-Soria,和C. Abanto-Rodriguez。2016.Estimación森林生态系统的碳水化合物año de instalación en el departmento de Huánuco。叶形线Amazonica 25(1): 45 - 54。https://doi.org/10.24841/fa.v25i1.382
Tovar, C., A. C. Seijmonsbergen, J. F. Duivenvoorden. 2013。监测山区土地利用和土地覆盖变化:以秘鲁安第斯山脉的哈尔卡草原为例。景观与城市规划112:40-49。https://doi.org/10.1016/j.landurbplan.2012.12.003
Vallet, A., B. Locatelli, H. Levrel, C. B. Pérez, P. Imbach, N. E. Carmona, R. Manlay和J. Oszwald. 2016。哥斯达黎加Reventazón森林过渡期间生态系统服务的动态。《公共科学图书馆•综合》11 (7):e0158615。https://doi.org/10.1371/journal.pone.0158615
范戴克,A. I. J. M.和R. J.基南。2007。种植的森林和水的透视。森林生态与管理51(2):1-9。https://doi.org/10.1016/j.foreco.2007.06.010
韦布洛娃,H., B.洛伊卡,T. P.丈夫,M. E. C.赞斯,P.云顿,A.罗洛,M.卡劳索瓦。2014。秘鲁亚马逊圣亚历杭德罗可可农林的树木多样性。农林复合经营系统88(6):1101 - 1115。https://doi.org/10.1007/s10457-013-9654-5
Vélez Quesquén, M. A. M.和J. I. Saavedra Chávez。2019.帕盖班巴,奎罗科托,乔塔,卡哈马卡,Perú, 2018。(鞘翅目:金龟子科:金龟子科)论文。秘鲁兰巴耶克国立佩德罗·鲁伊斯·加洛大学。https://hdl.handle.net/20.500.12893/5511
维贾伊,V., C. D.里德,M.范纳,C. N.詹金斯和S. L.皮姆。2018。秘鲁亚马逊地区油棕的扩张带来了砍伐森林的风险。环境研究通讯13(11):114010。https://doi.org/10.1088/1748-9326/aae540
维拉尔·卡贝扎(Villar Cabeza), M. Á。,J. E. Cuellar Bautista, and S. L. Valentin Castañeda. 2014. Valoración técnica, económica y ambiental de tres sistemas de silvopasturas en la región Cajamarca. Instituto Nacional de Innovación Agraria, Lima, Peru.http://repositorio.inia.gob.pe/handle/20.500.12955/376
Villogas Ventura, K. E. 2013。可可森林系统碳化铝(SAF)Theobroma可可produccion l .)。论文。国家农业大学,秘鲁廷戈María。http://repositorio.unas.edu.pe/handle/UNAS/163
Vuille, M., M. Carey, C. Huggel, W. Buytaert, A. Rabatel, D. Jacobsen, A. Soruco, M. Villacis, C. Yarleque, O. E. Timm, T. Condom, N. Salzmann, J.-E。Sicart》2018。热带安第斯山脉冰雪的迅速减少——未来的影响、不确定性和挑战。地球科学评论176:195 - 213。https://doi.org/10.1016/j.earscirev.2017.09.019
Vuille, M., B. Francou, P. Wagnon, I. Juen, G. Kaser, B. G. Mark, R. S. Bradley. 2008。beplay竞技气候变化与热带安第斯冰川:过去、现在和未来。地球科学评论89(3 - 4):79 - 96。https://doi.org/10.1016/j.earscirev.2008.04.002
Walentowski, H., S. Heinrichs, S. Hohnwald, A. Wiegand, H. Heinen, M. thicth, O. A. Gamarra Torres, A. B. Sabogal和S. Zerbe. 2018。Pomacochas盆地(亚马孙,秘鲁北部)退化地点的植被演替——森林恢复的生态选择。可持续性10(3):609。https://doi.org/10.3390/su10030609
Whaley, o.q, d.g. Beresford-Jones, W. Milliken, A. orelana, A. Smyk,和J. Leguía。2010.秘鲁南部干旱森林恢复和可持续管理的生态系统方法。丘公报65(4):613 - 641。https://doi.org/10.1007/s12225-010-9235-y
惠特沃思,A., R.唐尼,R.冯·梅,J.维拉坎帕和R.麦克劳德。2016.一个再生的雨林能提供多少潜在的生物多样性和保护价值?秘鲁亚马逊的“最佳情况”方法。热带保护科学9(1):224-245。https://doi.org/10.1177/194008291600900112
威尔逊、S. J.舍尔哈斯、R.格劳、A. S.南尼和S.斯隆。2017。森林生态系统-服务转变:森林转变的生态层面。生态与社会22(4):38。https://doi.org/10.5751/ES-09615-220438
杨,K. R.和B. León。2007.沿安第斯山脉的林木线变化:空间模式和动态的影响。皇家学会哲学学报B 362(1478):263-272。https://doi.org/10.1098/rstb.2006.1986
Zamorano-Elgueta, C., J. m.r. Benayas, L. Cayuela, S. hanson和D. Armenteras. 2015。1985-2011年,智利南部的外来种植园取代了原生森林,并通过自然再生进行部分补偿。森林生态与管理35:10 -20。https://doi.org/10.1016/j.foreco.2015.02.025
扎瓦拉,W., E.梅里诺,P. Peláez。2018.可可种植系统,森林种植系统,以及碳的种植系统。Scientia Agropecuaria 9(4): 493 - 501。https://doi.org/10.17268/sci.agropecu.2018.04.04

图1

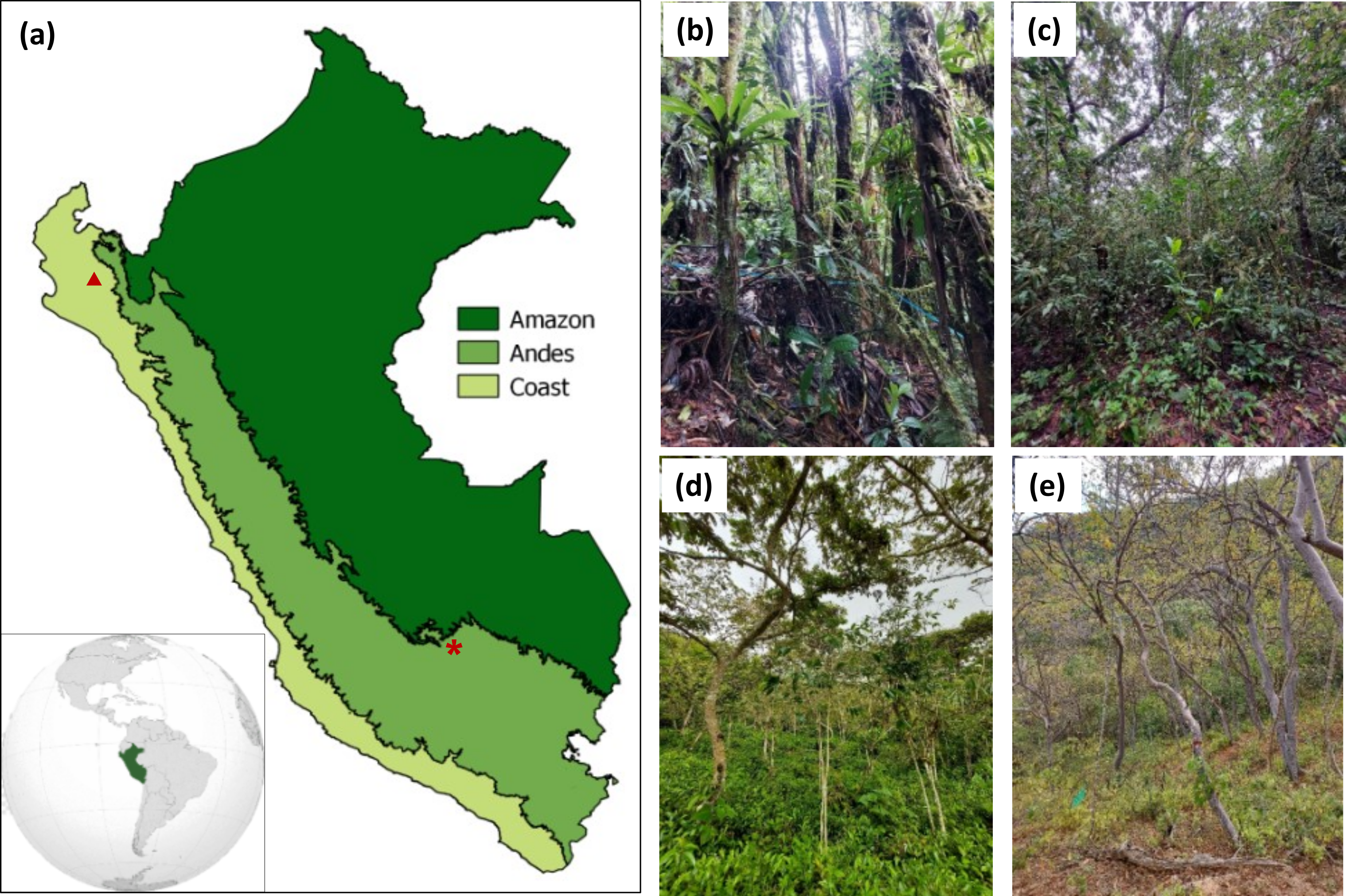
图1所示。(A)秘鲁沿海、安第斯和亚马逊地区的地图。照片显示的是成熟森林(B)、茶园之后的次生林再生(C)和茶叶农林复合系统(D),所有这些都位于库斯科地区的华奥帕塔区(在图a中用星号表示)。(E)位于秘鲁西北部皮乌拉地区Mangamanguilla私人保护区的沿海干燥次生林照片(在图a中用三角形表示)。
图2
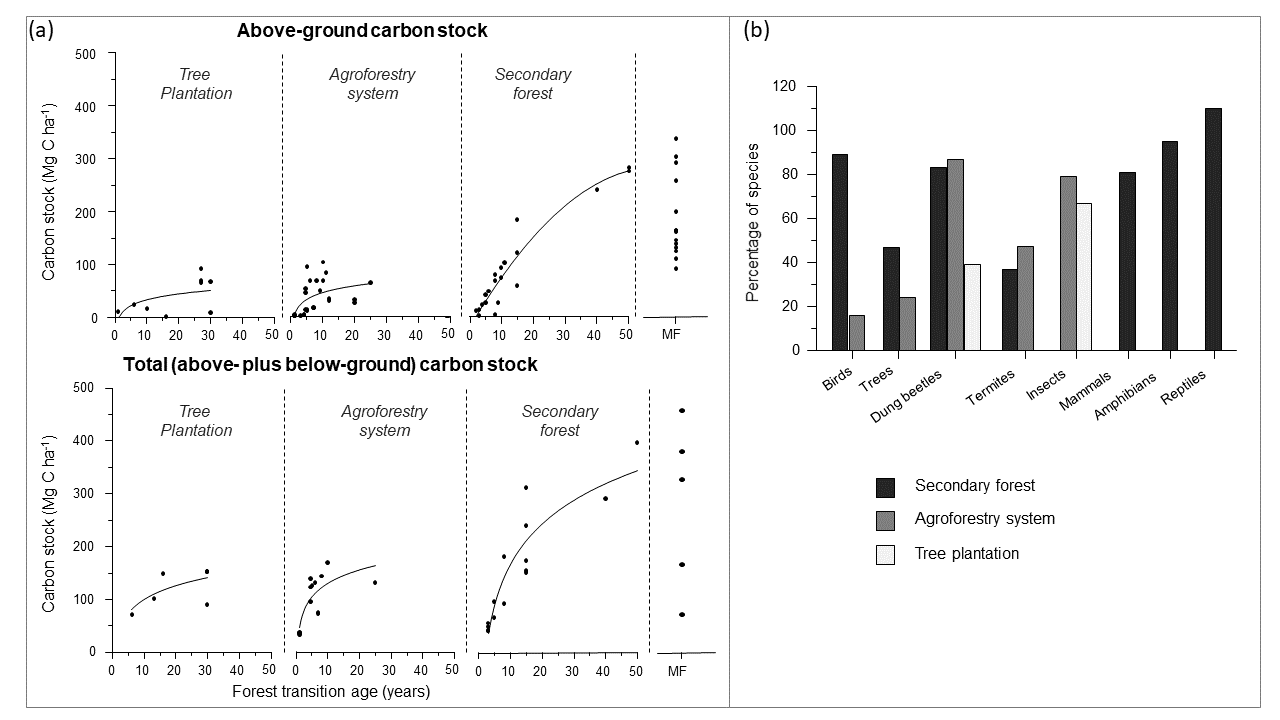
图2所示。(a)亚马逊地区次生林和两种人工辅助的毁林后再生林(人工林和农用林业系统)的碳储量和(b)生物多样性价值。在图2b中,物种的百分比表示与成熟森林基线相比的百分比。MF:成熟的森林。这些数字所用的数据载于表A2。
图3
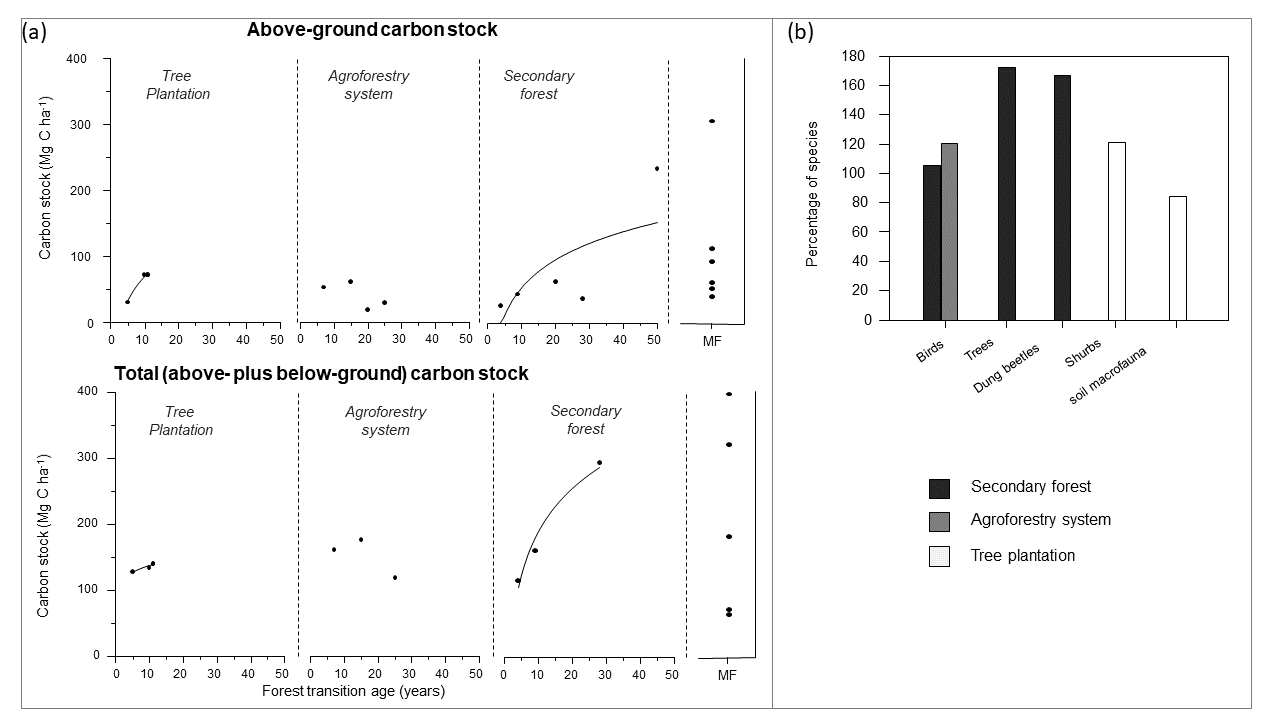
图3。安第斯区域次生林和两种人工辅助的毁林后再生林(人工林和农用林业系统)的碳储量和(b)生物多样性价值。在图3b中,物种的百分比表示与成熟森林基线相比的百分比。MF:成熟的森林。这些数字所用的数据载于表A2。
图4
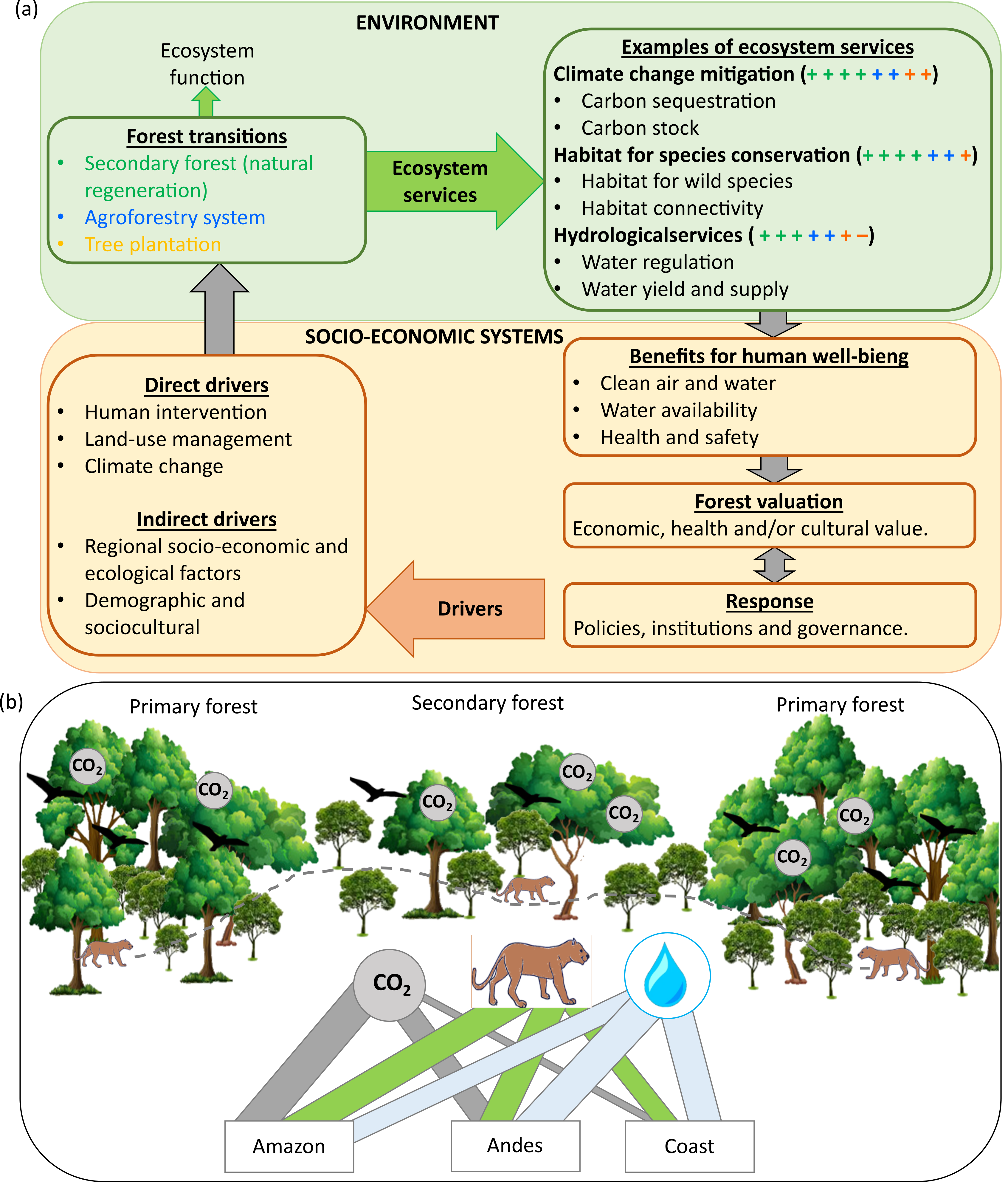
图4。(A)显示森林转变、其相对生态结果及其驱动因素之间关系的框架,将环境和社会经济系统联系起来。森林过渡的定义是森林面积的长期下降,被森林恢复取代。“+”和“−”表示根据林转换类型期望返回的每个服务的潜在数量。(B)亚马逊、安第斯和沿海地区次生林提供的关键生态系统服务的相对重要性。