






以下是引用这篇文章的既定格式:
Schrenk, H., B. Magnússon, B. D. Sigurdsson和W. zu Castell. 2022。苏特西岛植物群落发育的系统分析。生态与社会27(1):35。摘要
基于我们最近开发的量化Gunderson和Holling适应周期的方法,我们对2000年至2018年Surtsey火山岛上的维管植物群落进行了整体分析。在研究期间,我们发现了一个完整的适应周期,它反映了系统从一个典型的先锋到草原群落的转变。我们的研究结果支持了一个假设,即繁殖的海鸥带来的营养是这种发展的主要驱动力。研究阶段包括第二个周期的开始,这与隐喻中描述的模式不同。事实上,这一周期的开发阶段被所有三个系统变量同时下降所打断。我们可以将这种现象追溯到2012年的一次严重干旱。此外,该方法还允许我们建立个体物种在成熟过程中的系统作用。介绍
分析生态系统变化的动力仍然是一项具有挑战性的工作。生态系统的特征是由物理和化学过程,以及居住在系统中的生物的相互作用,它们对特定环境的适应,以及后者的形成形成的。
生态系统发展的动力学已经被广泛研究,并提出了一些定性(Bossel 1992, Holling 2001)和定量描述(Odum 1983, Ulanowicz 1986)的框架。Gunderson和Holling的适应性周期隐喻(Gunderson和Holling 2002)是定性描述之一,这是他们的全社会论理论的核心。根据这个比喻,系统的发展是由三个综合的系统属性决定的:(1)系统对未来变化可用的潜力,(2)系统内部变量和过程之间的连通性,以及(3)系统在不可预测的扰动下的恢复能力。这三个属性的相互作用定义了四个不同程度的可预测性阶段。当遇到大量未开发的资源时,系统将开始利用可用的资源来发展。在这一时期,生态系统以所谓的r-战略家(r- strategies)为主,其特征是高散布能力和对栖息地的低需求(Pianka 1970)。按照这个术语,这个时期被称为”r步”或者说开发阶段。通常情况下,弹性是很高的,而潜力和连接仍需建立。随着时间的推移,该系统的内部组织水平提高,从而获得资源,例如,反映为新利基的开放(Odling-Smee等,1996年)。然而,专业化水平的提高导致刚性的增加,导致弹性的下降。按照生态学术语,这个阶段被称为”K步”或者说守恒阶段。它的特点是高度活跃和效率,导致高水平的潜力和连接。在一起,r- - -K-阶段包括一个巩固和可预测的时期。在某一时刻,一个触发会导致既定结构在所谓的破坏”Ω步。”资源正在释放,以前的连接已经断开。潜力和连通性大幅下降,而韧性却在增强。最终,动员过程启动了系统的α-阶段或重组阶段。在此期间,新的机会被利用,新的组件可能进入系统,而其他组件可能丢失。后两个阶段因此具有高度的随机性,允许创造性变化的机会。随着一组组件慢慢就位,系统开始进入另一个r阶段。根据这个比喻,这种生长与更新、巩固与创新之间的交替,允许一个复杂的系统反复适应不断变化的环境。
由于隐喻具有高度的通用性,它可以作为开发各种复杂系统的有价值的框架。然而,它也有其局限性。例如,被动地与外部可变性生活的系统以及预测和操纵可变性的系统可能永久地保持在单一阶段(冈德森和霍林2002年)。此外,内外因素的相互作用通常会导致单相的跳过或拉伸。尽管在直觉上是合理的,但由于量化潜在驱动变量的潜力、连通性和弹性的内在困难,适应周期迄今为止在很大程度上未能得到经验验证(例如,Burkhard等人2011年,Fath等人2015年)。在Castell和Schrenk 2020中,我们提出了一种量化适应周期的方法。该方法利用了信息论的抽象框架,具有普遍适用性,只需要关于系统的最基本信息,即其组件丰度数据的时间序列。这为应用开辟了广泛的可能性,因为为了尽量减少对观测系统的干扰,物种丰度计数仍然是生态系统评估最常用的方法。
我们的目标是根据Castell和Schrenk 2020引入的方法,对Surtsey岛上维管植物群落的发展进行整体分析。1963年,火山爆发在冰岛的一个近海岛屿Heimaey西南形成了一个岛屿。这个新形成的岛屿被命名为Surtsey,从那时起,它的地质和生物发展一直受到密切监测(Fridriksson 2005)。在它形成的两年后,第一种高等植物在岛上被观察到,尽管它还不能坚持(Fridriksson 2005)。到1985年,一个稳定的先锋植物群落在火山岛的南部发展起来,那里的地形躲避了冬季的巨浪,海鸥开始繁殖,带来了更多的种子和营养物质(Magnusson et al. 2014)。从那时起,维管植物群落经历了几个阶段的进化,为评估生态系统的发展提供了有价值的数据集。
通过分析2000 - 2018年在一个永久监测地块上的维管植物群落,可以识别出生态演替模式,反映了群落从早期拓荒者向稳定的草地生态系统的过渡。在研究期间,系统运行一个完整的适应周期,并开始进入另一个周期。我们的分析支持将繁殖的海鸥作为第一个系统崩溃的外部驱动因素的识别。第二个循环表明,环境条件的显著变化导致了隐喻的偏离。实际上,在第二个周期中,系统的开发阶段突然被所有三个系统变量同时下降所中断。这种现象可以追溯到一次严重的干旱。
方法
计算自适应周期
Castell和Schrenk 2020详细介绍了我们的自适应周期操作化方法。因此,在这一点上,我们只简要回顾一下基本思想。包括显式数学公式的技术说明可在附录1中找到。
考虑一个交互代理的系统。假设对于一组固定的时间点,每个代理的状态是已知的。我们称之为数据”丰富的数据,”虽然我们并不把自己限制在“富足”这个词的严格意义上。对数据的唯一要求是,它们至少隐含地捕获了代理之间交互模式的定性性质。就生态系统而言,这类数据可以是生物量或每个物种的个体数量。在经济体系中,资本可以量化公司的丰富性。我们现在假设,每一次有效的交互都会导致相互作用的主体之间的信息传递,并在相应的丰富数据的时间序列之间的相互依赖性中显示自己。在这些假设的基础上,我们使用Schreiber的转移熵(Schreiber 2000)来量化相互依赖性。来自agent的传递熵Y对代理X衡量代理结果的随机性X的丰富性可以用代理人的知识来解释Y的丰度。换句话说,我们能对未来了解多少X不仅知道过去的X但是过去Y吗?我们估计每一对有序代理之间的转移熵。通过将代理视为节点,将信息传输视为边,结果可以用加权有向图的形式表示。而不是整个时间序列,我们只基于一个特定的时间窗口进行网络估计。通过移动这个时间窗口,我们得到了一系列网络,反映了系统信息结构的时间发展。为了简单起见,我们将这些网络称为系统的网络”信息网络。”从一个节点传输的信息总量将用它的信息表示”流出”以及被传输到一个节点的信息量,也就是它的信息”流入。”
我们将潜力、连通性和弹性定义为系统信息网络的属性。因此,在该方法的第一步中获得的一系列网络作为确定三个系统变量的发展的基础,独立于系统的具体实例化。我们对潜能和关联性的定义是受到Ulanowicz的能力和优势概念的启发(Ulanowicz et al. 2009)。势是信息网络中平均熵的度量(比较Cover和Thomas 2006)。因此,它抓住了可用的变化潜力,换句话说,它的未来可能的选择范围。在信息网络中有许多等权边的情况下,它是高的。直观地说,多样化的相互作用结构反映在广泛的信息传递中,使系统能够根据环境变化转移其重点。
连通性度量一个节点的信息流出和另一个节点的信息流入之间的平均互信息,由网络中的边缘实现(比较Cover等人,2006年)。因此,它量化了组件之间的相互约束和依赖关系。在没有冗余的有效信息通道的情况下,它是高的。请注意,我们对潜力和连通性的测量都是根据总系统吞吐量(即所有信息传输的总和)进行缩放的。因此,这些措施随着整个系统活动的增长而增加。
我们对弹性的定义类似于已建立的谱图-连通性理论度量(Chung 1977)。连通性量化了无向图对扰动的脆弱性。更准确地说,它表明如何通过现有的替代路径来补偿图中单个边缘的损失。让我们回忆一下甘德森和霍林对弹性的定义”在系统改变控制行为的变量和过程之前,可以吸收的扰动的大小”(甘德森和霍林2002:28)。从这个意义上说,连接抓住了韧性的概念,作为一种衡量”安全网”系统的信息结构。我们把连通性的概念转移到有向图上。系统变量的数学定义见附录1。
R包QtAC(量化适应性周期)允许直接应用我们的方法(https://github.com/hannahschrenk/QtAC).Schrenk等人(2022)介绍了该软件包,并说明了其应用。我们使用的计算基础的R代码在附录2和3中提供。
案例研究的数据和参数:Surtsey
我们的分析基于在Surtsey南部一个10 x 10米的地块(地块1)收集的维管植物物种丰富度数据的时间序列(见图1 b, d;该地块是填沙熔岩片上海鸥筑巢点的一部分(图1 d)。在观察期间,可见12种不同的维管植物(见表1)。
1990年至2018年,农业研究所和冰岛自然历史研究所每两年在7月收集一次数据。继物种丰度之后,从2004年开始,研究人员统计了这块土地内和周围的鸥巢数量。由于岛上植物与繁殖鸟之间有很强的相互作用,因此我们将这个量以可变筑巢密度(ND)的形式纳入分析。有关数据收集和监测方法的更多信息,请参见Magnússon等人(2009)。降水数据来自Stórhöfði上的气象站,Stórhöfði是Vestmannaeyjar群岛的主岛,距离Surtsey约18公里(由冰岛气象局2019年1月提供,附录5)。我们将12种维管植物物种和筑巢密度作为一个联合复杂系统的组成部分。为了估计信息网络并计算潜力、连通性和弹性,我们使用了R包QtAC (https://github.com/hannahschrenk/QtAC).
按照附录1中的术语,我们选择一个固定的窗口大小为W = 6,因此,每个信息网络的估计是基于前12年。这个参数的选择总是在统计可靠性(窗口中有大量值)和结果的充分时间分辨率之间进行权衡。一般来说,我们不建议使用小于5个数据点的窗口大小(比较Schrenk et al. 2022)。在目前的情况下,选择窗口大小相对较小的有两个原因W = 6.首先,对于一个对应两年的数据点,结果的时间分辨率的平均程度和平滑程度随着窗口大小的增加而显著增加。第二,窗口大小越大,第一个估计网络的时间点越晚。在本案例研究中,我们要特别观察2004年候鸟到达的系统效应,因此有必要提前一段时间考虑系统。
我们选择了一个统一的历史长度K = l = 3.因为出于统计可靠性的考虑,我们对现有数据进行了插值,所以这个历史长度对应于两年的时间段(参见附录1)。这一选择基于对系统动态的评估。就它们的时代性而言,四年的历史长度似乎不合适。数据情况不允许有更好的分辨率。
只有当其显著性≤0.1时,才考虑估计的信息转移。我们不建议使用显著性> 0.1来保证估计的统计可靠性。
弹性是根据底层网络的最大边权归一化的(比较Schrenk et al. 2022)。因此,系统的弹性只取决于相对边缘权重,而不是被传输的信息总量。
我们将估计过程中添加的随机高斯噪声量设置为1e-20。这种选择是基于敏感性分析的。附录6显示了基于50次运行的系统变量的样本噪声水平的平均值和标准差。尽管电势和连通性在1e-20以下的噪声水平上变化不大(附录6 a, b),但当噪声水平高于该值时,标准差明显增加(附录6 c)。在某些年份,即使在较小的噪声水平下,复原力的标准差仍然相当大。这是因为弹性衡量的是局部脆弱性,而潜力和连通性是全球网络属性。因此,弹性对网络中是否存在单个边高度敏感。在这种情况下,最好是寻找高标准偏差的原因,并在系统分析中考虑到这些见解。
结果与讨论
为了确定一个具有代表性的系统变量的发展,我们考虑了我们的方法的50次运行的结果,参数设置在方法部分中指定。图2 (a)显示了这些运行的平均值和标准偏差。在电位和连通性都存在的情况下,标准偏差小到足以明确地识别各自系统变量的发展。然而,该系统的弹性在2000年、2002年、2006年、2010年和2012年显示出相对较高的标准偏差。图2 (b)以散点图的形式显示了值的分布。可以观察到,弹性在2002年是均匀分布的,而在其他年份是围绕两个值分组的。后者可以追溯到单个边的(不)存在,估计其显著性接近于0.1的阈值水平。因此,有时边缘被包含在网络中,而在其他运行中被排除在外。我们选择一个代表大多数结果的值作为系统的弹性。在均匀分布(2002年)的情况下,我们选择一个平均值作为代表性的弹性。 However, interpretation must be taken with care for this value.
图1 (a, c)说明了系统系统变量的代表性发展。三维情节从两个不同的角度展示。颜色突出显示了已识别的适应周期阶段,绿色表示稳定阶段,红色表示衰退阶段,橙色表示过渡阶段。对研究地块上维管植物群落的分析显示出四个明显的阶段。
我们的观察期开始于r-阶段。通常情况下,潜力和联系在这一时期增加了。整体复原力也有所增强,这是相当不寻常的。
一个清晰的K-阶段可确定为2004年至2006年。连通性进一步增强,同时适应力大幅下降。通常是晚期K-phase时,电位开始下降。
这一阶段因2006年开始的一次系统故障而中断。由于数据是以两年为基础提供的,我们决定将随后的两个随机阶段合并为一个Ω/α-阶段从2006年到2010年。它开始于潜力和连通性的特征下降与弹性的同时增加。从2008年到2010年,潜力和连通性开始再次增加,同时适应力也在增加,这是典型的α步。
从2010年到2012年,我们看到了第二种情况r-阶段,表现为潜力和连通性的增加,伴随着韧性的下降。2012年,系统的开发阶段突然中断,三个系统变量同时下降。在那一年之后,这些变量继续在其当前值附近振荡。它们之间的相互作用并没有表现出适应周期阶段的典型特征。这种非典型发展通常暗示着外部驱动因素,从本质上改变了系统的特征。
总的来说,在研究期间,该系统运行了一个完整的周期,并开始进入第二个周期。从2000年到2006年,一个经典r / K的开发和保护阶段先于四年Ω/α-释放和重组阶段。虽然刚德森和霍林预料到Ω/α相比较而言非常短r / K-阶段(Gunderson and Holling 2002),我们得出结论,在自然系统中,重新定位和重组的周期可能要长得多。
2010年,该系统进入了一个新的周期,建立了潜力和连接,直到2012年。有趣的是,预期的进一步增长在2012年中断了。这种偏差表明基本环境条件发生了变化。事实上,2012年是一个特别的干旱年。
附录7(图a)演示了在不同窗口大小下的整体模式。对于w = 5和w = 7的窗口大小,研究期间的两个适应周期之间的过渡是清晰可见的。特别是在第二开发阶段,潜能和连通性的突然下降是时间稳定的。请注意,在第一个周期中连接的分解显示了从2006年到2010年窗口大小增加的时间转移。我们将在稍后的时间回到这个现象。
重要的是要记住,我们不能也不应该期望任何真正的系统完全遵循适应周期的理想模式。这个比喻应该被看作是一个系统的基线开发的描述,它可以被用作对给定实例的详细分析的背景。特别是,偏离理想的情况允许我们识别系统开发中的特殊性,这些特殊性是由于在具体情况中给定的特定交互模式造成的。
第一个r- - -K -阶段:鸟儿的到来
让我们首先考虑2000-2004年期间的物种组成。能够在恶劣条件下生存的健壮物种仍然占优势。Honckenya peploides而且Leymus arenarius,连同Cerastium fontanum构成了超过80%的土地覆盖,所有这些都表明了最早的植物在Surtsey上的殖民(Magnússon et al. 2014)。在这一时期的末期,这些早期殖民者的贡献下降到不到50%。2004年,草种Poa pratensis成为主导。这种多年生草形成了茂密的草席,在竞争中胜过了一年生的群落成员,如植物而且Sagina procumbens.植物群落的组成表现出从早期拓荒者向草原群落的明显过渡。成立已久的Honckenya与Leymus(Fridriksson 1992),在稳定的环境条件下,开始让位于具有更高竞争能力的物种。这种发展与土壤中养分含量的增加有关,这可以追溯到岛上繁殖的海鸟的数量的增长,它们为它们的幼鸟从海洋带食物到陆地(Magnusson et al. 2009)。在此期间,两个新的殖民者在这块土地上定居下来:羊茅属rubra在2002年Tripleurospermum maritimassp。maritima(同义词洋甘菊maritima)。
在信息传递的网络中,我们洞察到植物群落的内部功能(见图3)。新物种的到来清楚地反映在网络结构中。在2002年,Tripleurospermum maritimassp。maritima在信息网络中占据了中心位置(图3 a, b)。两年后,在2004年,同样的现象发生在羊茅属rubra(图3 . c)Leymus而且Poa pratensis,f . rubra通过地面上和地下广泛的横向铺展形成致密的垫子(Magnusson et al. 2009)。同一年,随着第一个海鸥巢被发现在地块(图4),筑巢密度移动到网络的中心位置。图5 (a, b)说明了这些组件的信息输出和流入的突然增加。虽然一个物种的信息流出可以解释为其他物种对其发展的反应程度,但我们把信息流入理解为一个主动适应环境的过程。在这两个新殖民者的案例中,2004年的信息流入占主导地位,反映了他们到达时的“活跃”地位。附录8和9演示了观测到的现象在不同窗口大小下的稳定性。
在2004年至2006年期间,这一趋势在社区组成方面有所加强。Honckenya peploides,Cerastium fontanum,Stellaria媒体,植物数量进一步减少,而Poa pratensis而且Leymus arenarius加强他们的统治地位。引发这种变化的一个合理的原因可能是海鸥在岛上这个地区繁殖多年的累积效应。鸟类既是营养的提供者(特别是氮),也是种子的来源(Magnusson et al. 2009, 2014)。因此,它们改变了系统所有组件的环境条件。
在该系统的信息网络中也可以清楚地观察到前几年的这种发展的加强(图3d)。Tripleurospermum maritimassp。maritima,羊茅属rubra,与筑巢密度紧密相连,构筑了网络的中心点。的信息流入如图5 (b)所示羊茅属rubra已进一步增加。网络的另一个中心位置被蒲公英这部电影于2006年进入剧情。这也反映在2006年信息流入和流出的增加中,见图5 (d)。
与2004年相比,信息网络的整体结构已经从许多权重相当均等的边变成了权重更少、权重更不均的边。这种拓扑的变化反映了发生在系统结构内部的聚焦,并负责增加连通性和降低电位。从生态学的角度来看,这一时期末期的极低恢复力可以进一步解释为仍然普遍存在的、僵硬的先驱者联想Honckenya而且Leymus.这些联系现在正随着人口的下降而慢慢消失Honckenya.
第一个分类:从先锋到多年生植物
在2006年至2010年期间,群落组成没有显著变化。该地块被较高级的竞争性物种所主导Poa pratensis而且Leymus.在此期间,两种植物一起提供了几乎所有的地表覆盖(见图1b)。然而,之前的重组最终导致了系统的崩溃。从2006年到2010年,网络结构发生了强烈的变化(图6)Ω/α-阶段的复杂系统。为了适应不断变化的环境条件,系统利用了不同的机会”试错法”模式。在2008年,Tripleurospermum maritimassp。maritima不再是紧密相连的三角关系的一部分羊茅属rubra和筑巢密度(图6 a)。蒲公英与后两者建立了密切的联系。从图5 (a)可以看出,与此同时,筑巢密度的信息流出占主导地位,捕捉到了种鸟对植被的强烈影响。这一现象可以解释为,第二演替波的特化和高要求的物种依赖于鸟类的养分输入。此外,还必须考虑鸟类干扰的影响,这可能使新物种得以殖民和扩张。
回想一下在不同窗口大小下系统崩溃的时间转移(附录7)。这种现象提示了这样一个事实,即崩溃的某些原因可以追溯到时间窗口的开始。更确切地说,我们可能会看到1996年以后社区组成的根本改组的后期影响。因此,在某些物种的信息传递中应该可以观察到类似的模式。举个例子,考虑一下信息流入和流出是如何突然下降的Puccinellia distans由2006年改为2010年,窗口大小增加(附录10)。的确,Puccinellia是一种在1996年达到丰产高峰,之后逐渐衰落的植物。
在2010年,网络稳定下来,呈现出一组相当平衡的边缘,熵流分布更加均匀。这表示结束Ω/α阶段和一个新的增长时期的开始。考虑到物种的信息流,未来的趋势出现了。虽然总信息流的嵌套密度和羊茅属rubra强势下滑,信息流泛滥Stellaria媒体开始增加(图5 a-c)。此外,信息网络显示出日益重要的地位Tripleurospermum maritimassp。maritima(图6 b)。在系统水平上,2008 - 2010年间,积累的养分和新建立的植物定植剂导致了潜力的增加。
在2006年和2010年期间,信息网络的激进重组在组件特征中心性的发展中特别明显(比较Csardi和Nepusz 2006年)。简单地说,在网络中,具有高特征中心性的节点与其他节点是强连接的,而这些节点本身也是强连接的。因此,特征中心性抓住了”重要性”网络中节点的。图7以热图的形式展示了2000 - 2018年信息网络中各成分的特征中心性。很明显,在2008年到2010年间,情况出现了转机。虽然筑巢密度的特征中心性和羊茅属rubra强烈下降(参见它们在网络中的突然边缘位置),草的特征中心植物而且Puccinellia distans大大增加。此外,正如在信息网络中已经注意到的,Stellaria媒体突然占据了中心位置。
第二个周期:计划的失败
在2010年到2012年之间,许多物种的数量增加了Poa pratensis,Leymus arenarius,Stellaria媒体.在2012年的系统信息网络中,前两年的发展有所加强(图6 c)Tripleurospermum maritimassp。maritima而且Stellaria媒体占据了网络的中心位置。也蒲公英再次占据了中心位置。信息网络就是典型的例子”收缩”系统的行为r阶段,重点关注一些信息传输。这种行为导致了联系的增加和弹性的降低。
从2012年到2014年,系统的r-阶段突然被所有三个系统变量同时下降打断。系统组成也发生了剧烈的变化:几乎所有物种的丰度在2012 - 2014年间都在减少Honckenya而且Cerastium在地块上逐渐绝迹(图1 b)。这种现象可以追溯到在开发过程中,缺水干扰了系统。图8 (a)显示了在距离Surtsey岛18公里的Vestmanneyjar气象站测量的植被期的平均降雨量。2012年,降水量长期最低低于50毫米。虽然2007年的降水量相对较低,但植物以前从未表现出如此严重的干旱症状(B. Magnússon,个人沟通2020年10月)。可以看到,图8 (b)中显示的地块1(附录11)的初级生产总值在2012年最低。造成这种现象的一个原因可能是2012年的无雨时间比2007年更长,阵雨也更少,无法真正湿润土壤(B. D. Sigurdsson,个人沟通2020年11月)。此外,2012年鸟类数量的峰值(见图4b)可能为植物提供了额外的压力。更准确地说,高营养输入导致地下生长减少,增加了植物对干旱胁迫的敏感性。除了这些外部原因,我们将极端后果追溯到这样一个事实:干旱袭击了植物群落,而在2007年,该系统正处于其开发阶段Ω步。
虽然这个系统是在潮湿的气候条件和鸟类的适度干扰下形成的,但植物群落突然面临完全意想不到的条件。制定的计划α由于意外的干旱,这一阶段注定要失败。社区表现出的特征是,一个系统发现自己处于它未曾适应的条件中。在这种情况下,该系统是脆弱的,其恢复力极低(图1a)。2012年之后,该系统(暂时)必须投资于管理极端环境条件,而不是建立未来增长的潜力。我们顺便指出,在奇数年份没有采集数据。因此,系统的确切临界点不可能在一年的精度内确定。
考虑到这一时期的信息网络,人们可以观察到Stellaria在2012年至2014年之间占据了非常重要的位置(图6 c, d)。图5 (c)进一步说明了这一事实,并且强调Stellaria通过2016年信息流入的主导地位,彰显了美国在社区中的积极作用。这一现象在各自的信息网络中清晰可见(图9)Stellaria在此期间,变化可以得到生态上的支持。注意它是Stellaria只有在这一年中,通过死亡的植物材料释放的营养物质和利用意外的机会(图1b),它们会增加。作为一种短命的一年生植物,生长速度通常很高,能够迅速大量生产种子(Grime et al. 1988),Stellaria从环境扰动所提供的机会中获利。
适应性周期简介
自适应性周期的隐喻最初发表以来一直是争论的主题。对连通性、潜力和弹性这三个特征变量的解释出现了问题。还有一些问题是关于按照既定顺序要通过的四个阶段的严格程度。从系统科学的角度来看,复杂系统的识别总是包括观察者的视角。因此,特征变量不可能存在普遍的物理实例化,因为它们的解释必然受制于观察者将其视为系统的模式。然而,根据我们对一个系统的理解,即一组相互作用的代理,允许根据相互作用的特征来解释变量。将系统信息网络中的边缘看作通信通道,我们对连接的度量抓住了通信通道分布的不均匀性。一旦系统开始更多地利用某些交互,而忽略其他交互,连通性就会上升。类似地,电位捕捉系统在遇到前所未有的情况时可以利用的平均可变性。因此,较大的势能意味着系统具有自由度”通信结构。”最后,弹性度量网络在外部扰动下失去相互作用的敏感性。
自适应周期的比喻捕捉了这些变量之间相互关系的自然顺序,这些变量是由系统维持生存能力的努力所产生的。因此,在具体分析中观察到这种理想的偏差使我们能够以类似的方式更多地了解一个特定的系统,因为理论概念允许我们理解实验观察。从这个意义上说,在Surtsey数据中确定的第二个周期使我们得出结论,植物群落的环境发生了实质性的变化。同样,第一个完整的周期反映了植物群落为应对鸟类出现而预期的适应。
因此,没有必要假设适应周期的各个阶段都具有一定的关系持续时间。尽管这似乎是有说服力的期望Ω/α-相要比r / K-阶段,这样的结论从系统科学的角度来看是没有意义的。的持续时间r / K-阶段,例如,反映了环境条件的可预测性和整体稳定性。在Surtsey的研究中,在2012年的异常扰动之后,地块1中的植物群落似乎需要更多的时间才能稳定下来。最后,适应生活需要并暗示随机性的存在,这是人类的主要特征之一α步。
结论
我们对2000年至2018年间Surtsey火山岛上维管植物群落的发展进行了全面分析。基于我们分析系统开发的抽象方法,一个精细化的分析揭示了在不同的环境条件下,某些植物作为变化的驱动因素的作用。在研究期间,我们已经能够确定两个适应周期。2006年的第一次崩溃可以追溯到该系统在前几年经历的社区组成的巨大变化。事实上,信息传递网络反映了这种从拓荒者到古典草原群落的转变。因此,我们的网络分析支持了这样一个假设:繁殖的海鸥将大量营养物质带入植物群落是这一变化过程的主要驱动因素。随着草类在植物群落中占据主导地位,另一个周期在2010年左右开始。不典型的是,开发阶段被所有三个变量的突然下降打断。在接下来的几年里,变量振荡,没有表现出适应周期的特定阶段的特征。我们将这一现象追溯到2012年的一次严重干旱。
人们对苏特西岛维管植物群落的发展有相当的了解。这些个体物种之间的相互作用以及它们应对环境因素的能力都得到了很好的研究。因此,Surtsey岛上的植物群落为研究适应性周期隐喻在分析生命系统中的适用性提供了一个理想的场景。将系统的发展与自适应周期给出的理想场景进行比较,偏差提供了对内部动态驱动的洞察。案例研究证明了该方法分析较少研究的系统和揭示变化模式的能力。这种基于信息网络的方法为揭示系统的交互结构提供了基础,从而揭示了单个组件的系统功能。
由于这种方法也可以应用于比较不同的系统时,对Surtsey上生命的观察数据库为更好地理解新形成的岛屿上生命的发展提供了进一步的机会。岛上分布着几个永久性的监测地块,暴露在不同的土壤、天气和环境条件下。特别是,海鸟和海豹的影响水平在整个岛屿上差异很大(Magnússon等人,2020年)。按照适应周期的比喻来比较这些不同的植物系统,可能会为植物群落在开发新的资源储备时的适应策略提供新的见解。
除了生态方面的见解,这个案例研究为我们的方法的可能性和局限性提供了一个有价值的测试场景。结果表明,对系统各组成部分及其相互作用有一定的背景知识可以用来为参数找到合理的选择。然而,参数选择应该通过敏感性分析来支持。
考虑到具体的参数,在估计传递熵时应特别注意添加随机高斯噪声的水平。如果估计导致系统变量的稳定值,则可以找到适当的噪声水平。然而,目前的研究表明,弹性,作为一个地方的衡量标准,仍然可以保持不稳定。在这种情况下,应查明原因,并应谨慎解释相应的弹性值。估计过程中的另一个关键参数是由变量的历史长度给出的。相应组件的背景知识变得尤为重要。如果没有这些知识,可以使用已经建立的方法来估计这个参数(参见示例Lizier 2014)。窗口大小的选择总是在统计可靠性和结果的时间分辨率之间进行权衡。选择了窗口大小后,我们建议验证在略微变化的窗口大小下结果的稳定性。
数据可用性
支持本研究结果的数据作为本文的补充材料提供。它们包括物种丰度数据(附录4)、降水数据(附录5)以及初级生产总量数据(附录11)。本文正文中用于生成图形的R代码见附录2和附录3(附录2.pdf,附录3.md)。所有使用的功能都在R包QtAC (https://github.com/hannahschrenk/QtAC)中提供。
文献引用
1992年。真实结构过程描述是理解生态系统及其发展的基础。生态模拟63(1):261-276。https://doi.org/10.1016/0304 - 3800 (92) 90072 - m
伯克哈德,B. D.法斯和F. Müller。2011.适应适应周期:生态系统特性和服务发展的假设。生态模拟222(16):2878-2890。https://doi.org/10.1016/j.ecolmodel.2011.05.016
Castell, W.和H. Schrenk. 2020。计算自适应周期。科学报告2020(10):18175。https://doi.org/10.1038/s41598-020-74888-y
钟福荣。1997。谱图理论。CBMS数学区域会议系列。美国数学学会,普罗维登斯,罗德岛,美国。https://doi.org/10.1090/cbms/092
T. M.和J. A.托马斯的封面,2006年。信息论的要素。Wiley-Interscience,霍博肯,美国新泽西州。
Csardi, G.和T. Nepusz. 2006。复杂网络研究的igraph软件包。InterJournal。https://igraph.org
Fath, B. D., C. A. Dean和H. Katzmair. 2015。引导适应周期:一种管理社会系统弹性的方法。生态与社会20(2):24。https://doi.org/10.5751/ES-07467-200224
弗里德里克松,S. 1992。Surtsey的维管植物1981-1990。1992.Surtsey研究进展报告10:17-30。(在线)网址:https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.535.2177&rep=rep1&type=pdf
弗里德里克松,S. 2005。叙尔特塞岛。生态系统形成的。冰岛大学出版社,
格里姆,J. P., J. G.霍奇森,R.亨特,1988。比较植物生态学:英国常见物种的功能研究。施普林格,Dordrecht,荷兰。
甘德森,L. H.和C. S.霍林。2002。Panarchy:理解人类和自然系统的转变。美国华盛顿特区岛。
霍林,2001。理解经济、生态和社会系统的复杂性。生态系统4(5):390 - 405。https://doi.org/10.1007/s10021-001-0101-5
利兹尔,2014。JIDT:研究复杂系统动力学的信息理论工具包。机器人与人工智能前沿1(2)。https://doi.org/10.3389/frobt.2014.00011
Magnússon, B, G. A. Gudmundsson, S. Metúsalemsson, S. M. Granquist. 2020。海鸟和海豹是苏特西岛植物演替的驱动力。Surtsey研究14:115-130。https://doi.org/10.33112/surtsey.14.10
Magnússon, S. H. Magnússon, S. Fridriksson。2009。1999-2008年苏特西岛植物定植和演替的进展。Surtsey Research 12:57-76。
Magnússon, B. S. H. Magnússon, E. Ólafsson,和B. D. Sigurdsson. 2014。苏特西岛植物定植、演替与生态系统发展。Biogeosciences 11(19): 5521 - 5537。https://doi.org/10.5194/bg-11-5521-2014
欧德林-斯密,F. J. K. N.拉兰,M. W.费尔德曼,1996。利基市场建设。美国博物学家147(4):641-648。
欧达姆,1983。系统生态学:导论。威利,纽约,纽约,美国。
皮安卡,1970年。关于r-和k -选择。美国博物学家104(940):592-597。https://doi.org/10.1086/282697
Schreiber, T. 2000。测量信息传递。物理评论85(2):461-464。https://doi.org/10.1103/PhysRevLett.85.461
Schrenk, H., C. Garcia-Perez, N. Schreiber和W. zu Castell, 2022。QtAC:一个r包,用于在自适应周期隐喻的框架中分析复杂系统开发。生态模型466:109860。https://doi.org/10.1016/j.ecolmodel.2021.109860
乌拉诺维奇,1986。生长与发展:生态系统现象学。Springer-Verlag,纽约,纽约,美国。
Ulanowicz, R. E., S. J. Goerner, B. Lietaer, R. Gomez. 2009。量化可持续性:弹性、效率和信息论的回报。生态复杂性6(2):27-36。https://doi.org/10.1016/j.ecocom.2008.10.005

图1

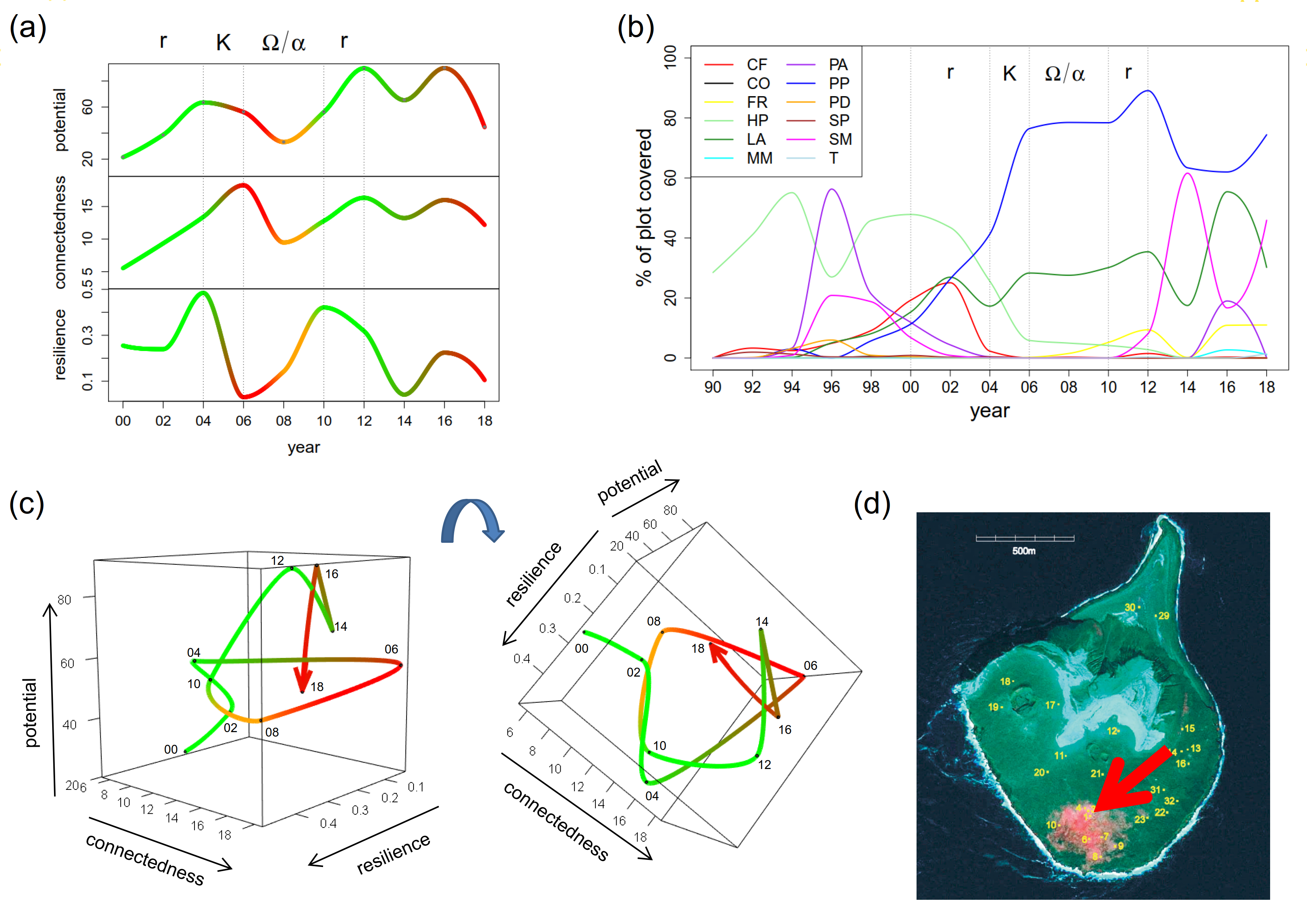
图1.(a) 2000 - 2018年维管植物系统的潜力、连通性和恢复力。(b) 1990 - 2018年第1地块物种丰度。(c)从两个角度分析2000 - 2018年系统适应周期的三维轨迹图。(d)地块1在Surtsey岛上的位置(Magnússon et al. 2009中图2的编辑版本)。
图2
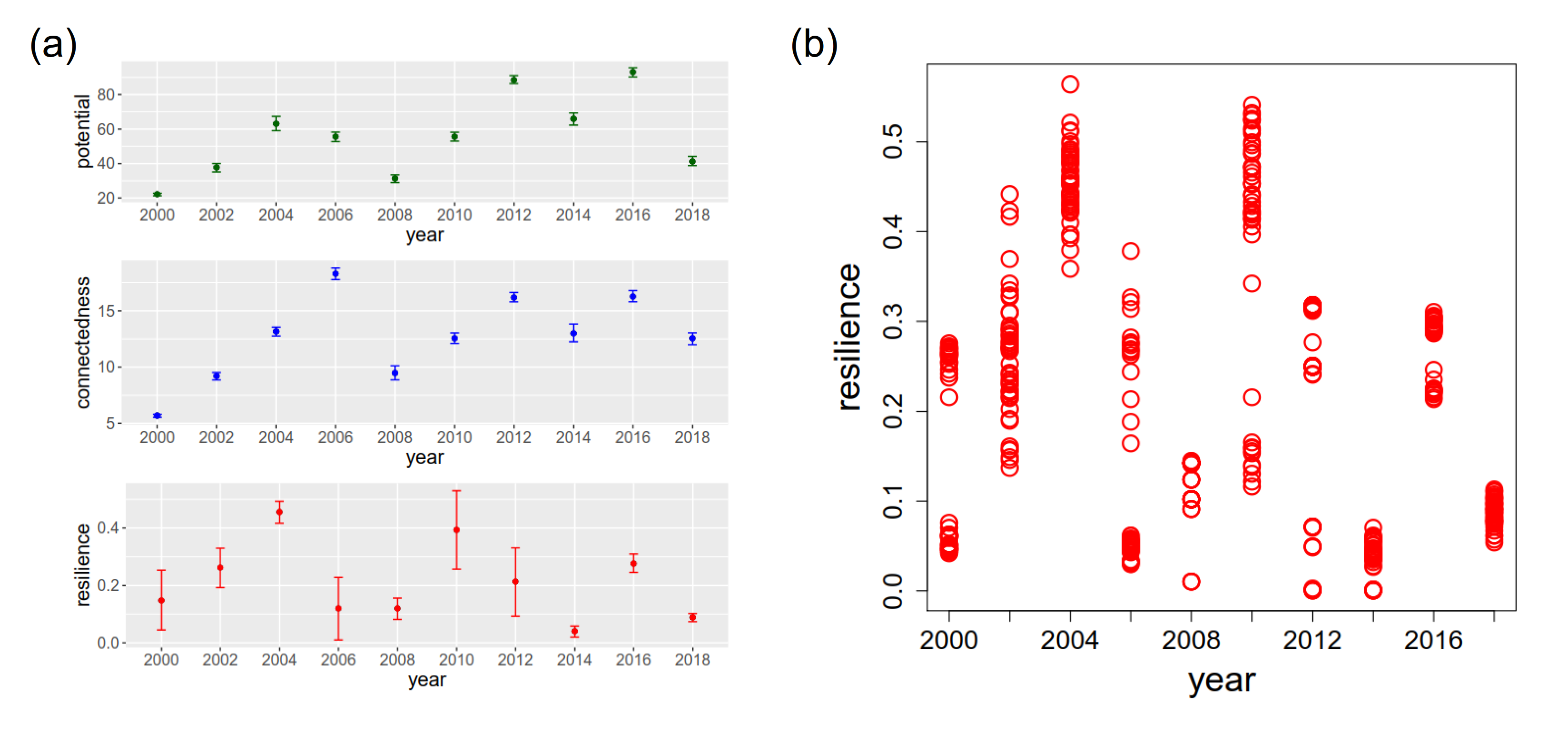
图2.(a) 2000 - 2018年维管植物系统的潜力、连通性和恢复力。(b) 1990 - 2018年第1地块物种丰度。(c)从两个角度分析2000 - 2018年系统适应周期的三维轨迹图。(d)地块1在Surtsey岛上的位置(Magnússon et al. 2009中图2的编辑版本)。
图3
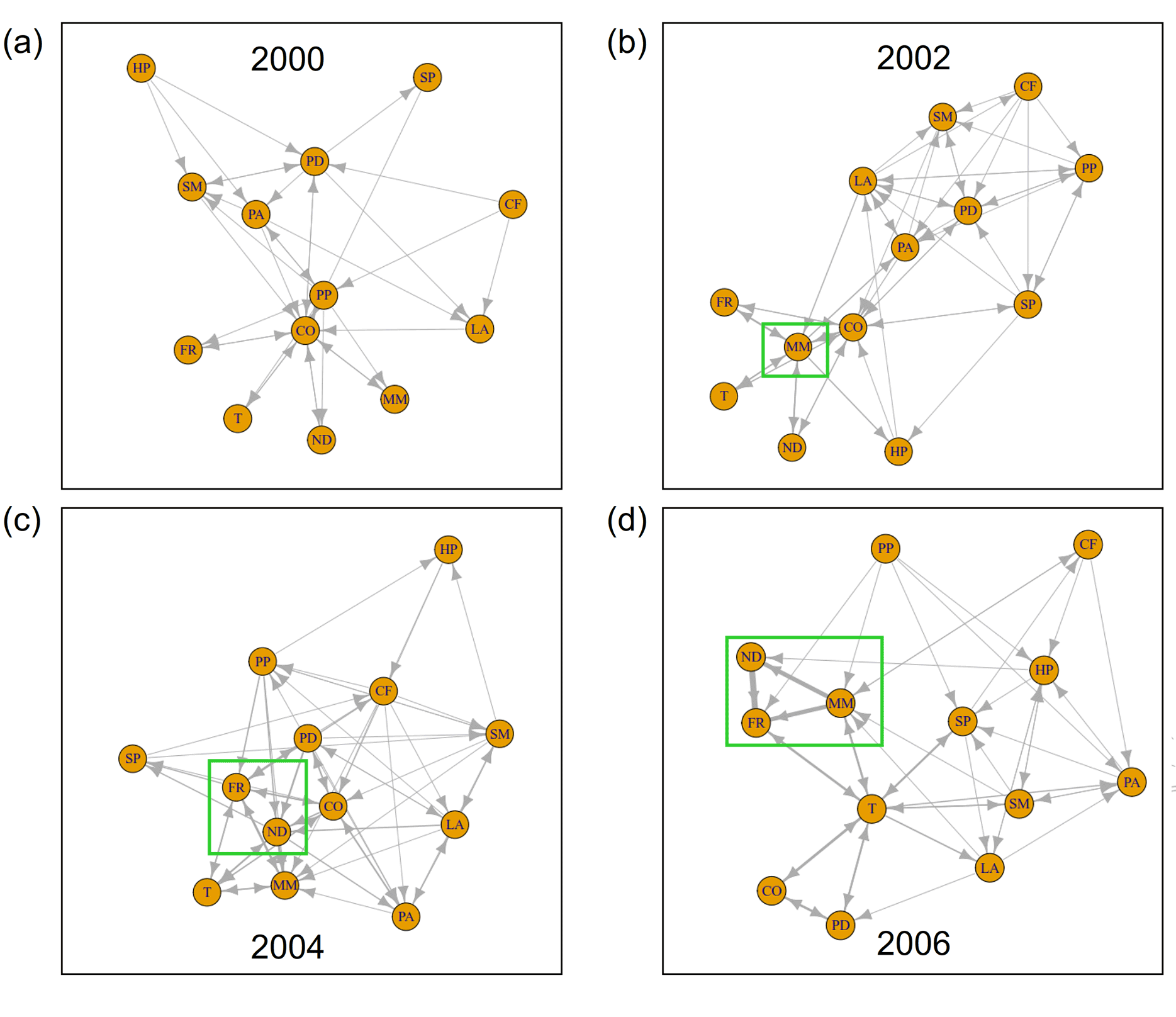
图3.2000年(a)、2002年(b)、2004年(c)和2006年(d)维管植物群落信息网络。边的宽度表示两个分量之间信息传递的强度。物种缩写在表1中展开。
图4
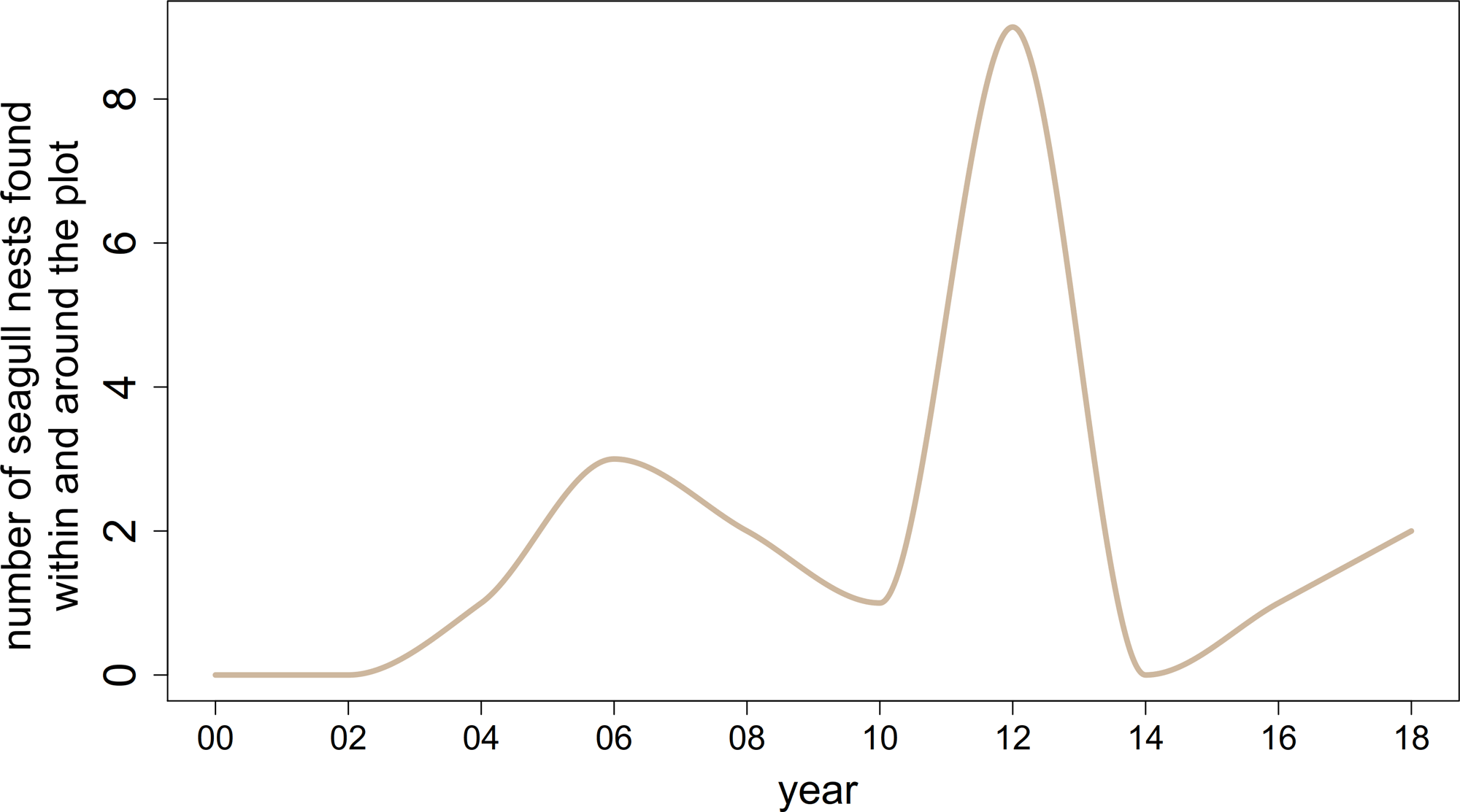
图4.2000年至2018年在地块1(面积1000平方米)内及其周围记录的巢穴数量。请注意,2004年以前没有统计过鸟巢。
图5
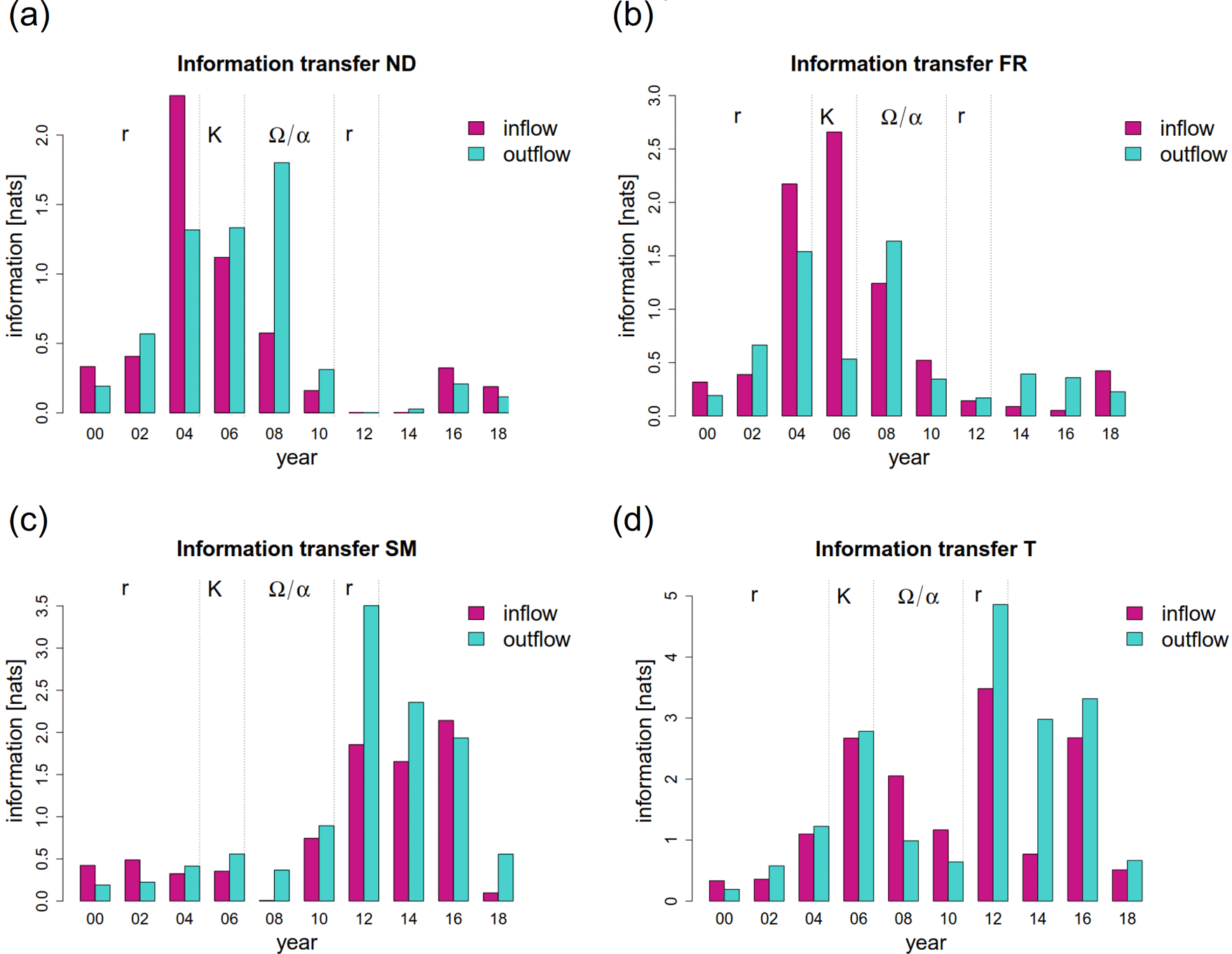
图5.信息流入和流出(a)嵌套密度(b)羊茅属rubra, (c)Stellaria媒体,及(d)蒲公英在2000年到2018年之间。信息以nats(自然信息单位)表示。
图6
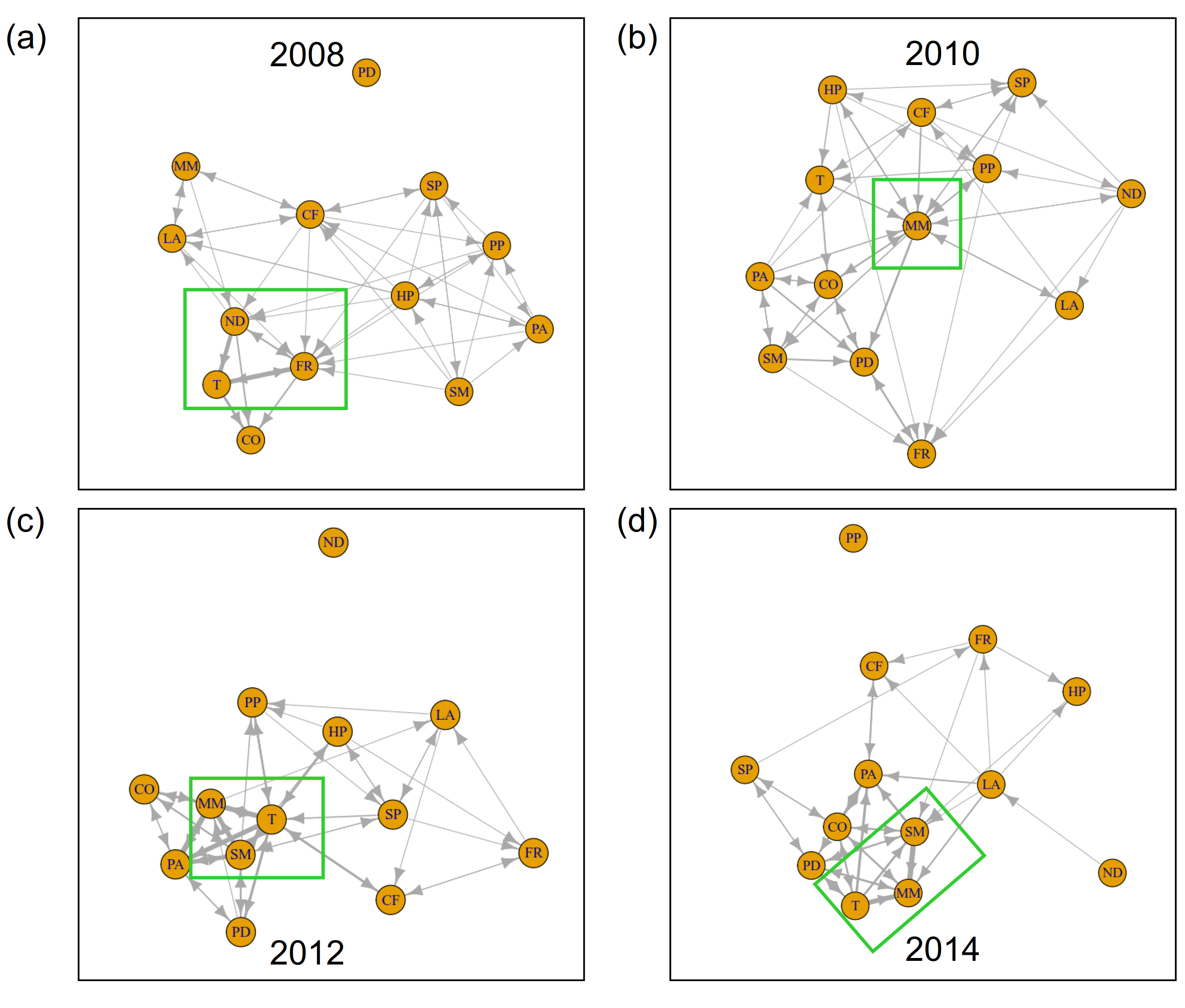
图6.(a) 2008年、(b) 2010年、(c) 2012年和(d) 2014年维管植物群落信息网络。边的宽度表示两个分量之间信息传递的强度。物种缩写在表1中展开。
图7
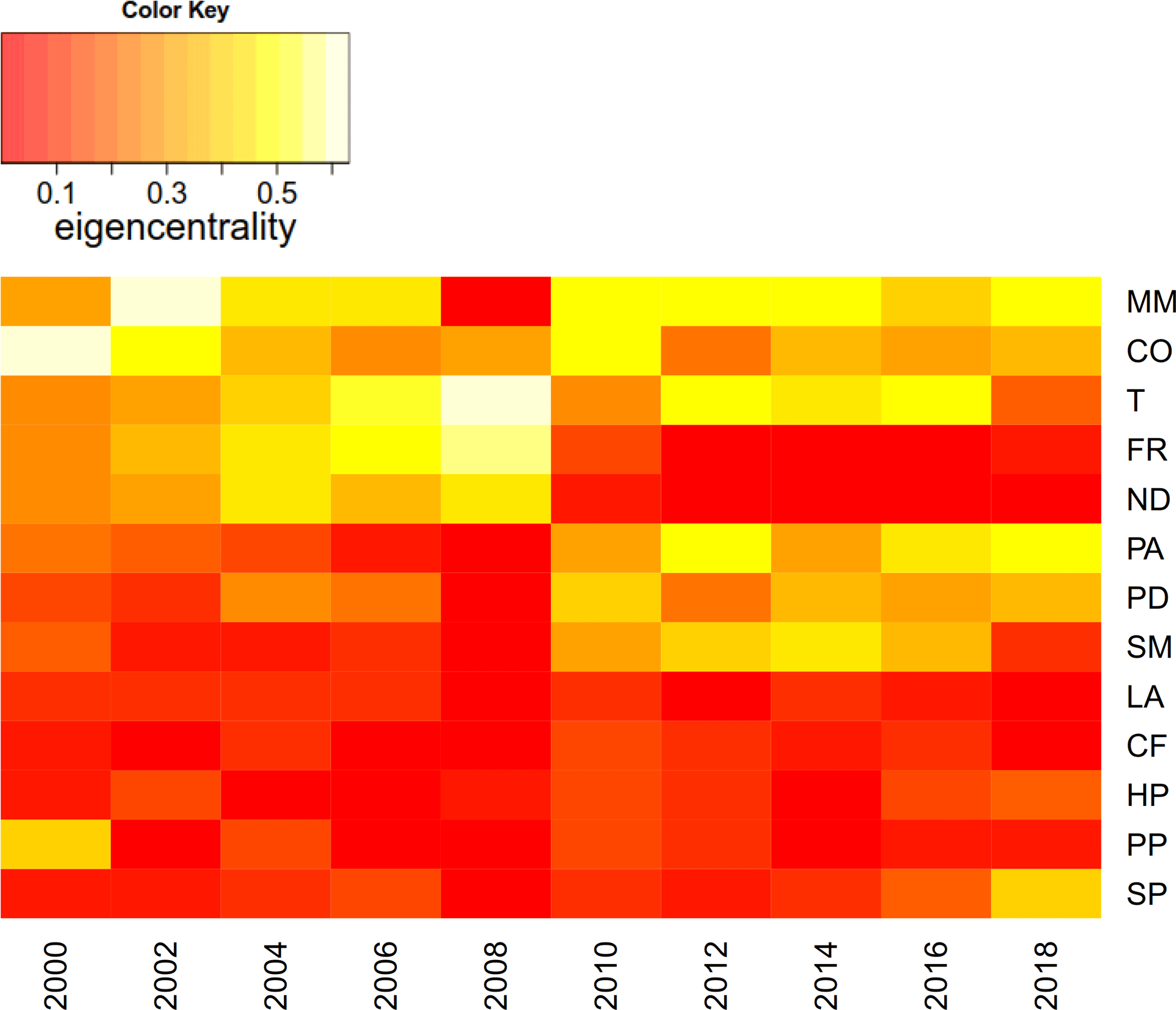
图7.2000 - 2018年信息网络中系统构件特征中心度热图。物种缩写在表1中展开。
图8
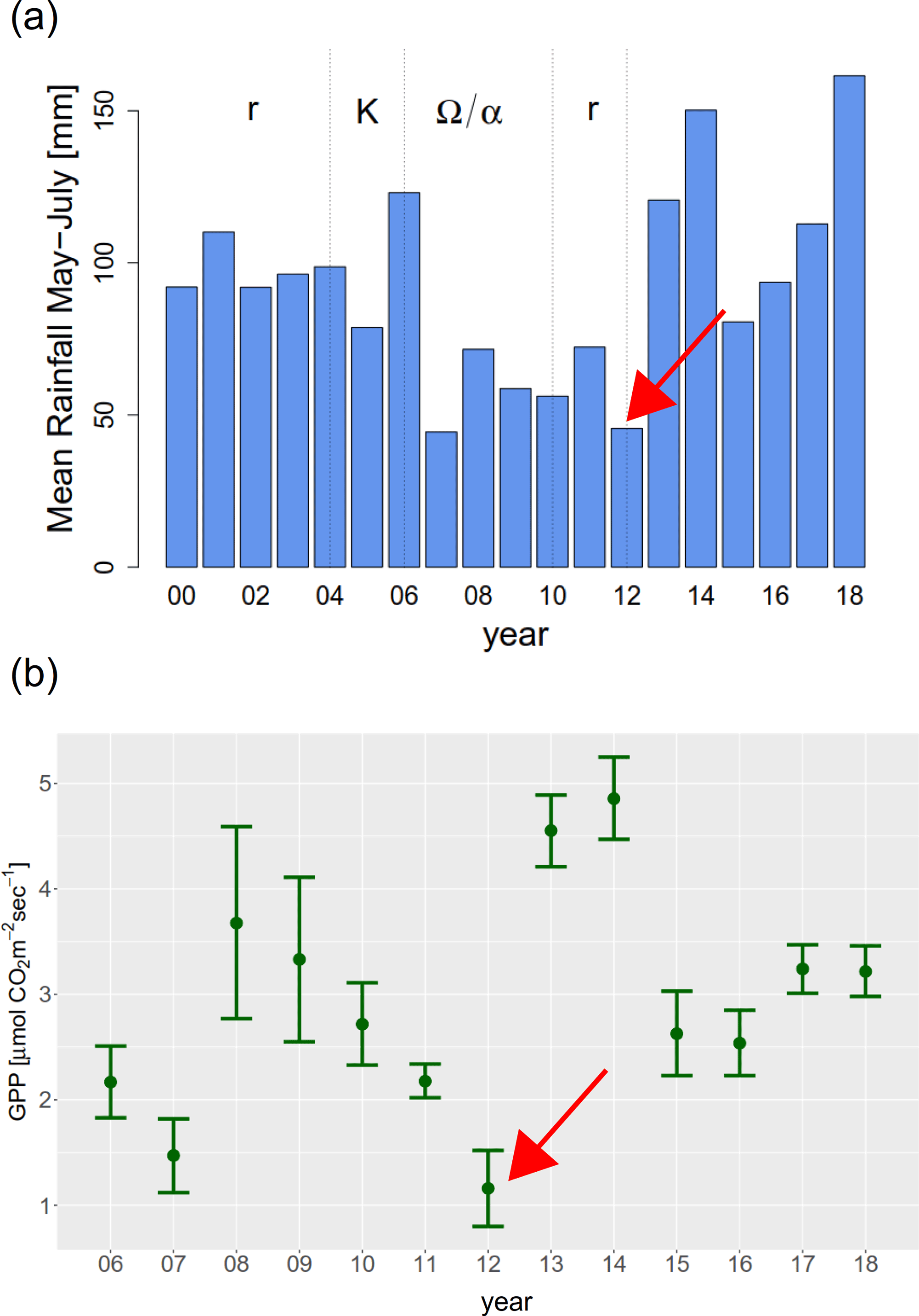
图8.(a) Vestmanneyar气象站测量的2000年至2018年5月至7月的平均降雨量。(b) 2006 - 2018年第1地块光饱和生态系统光合速率(GPP)±SE。GPP的测量由EGM-4便携式气体分析仪完成,并在透明室和暗室分别重复测量了4次通量。2016年的数值不是直接测量,而是通过地块1归一化植被差异指数的重复测量得出的。
图9
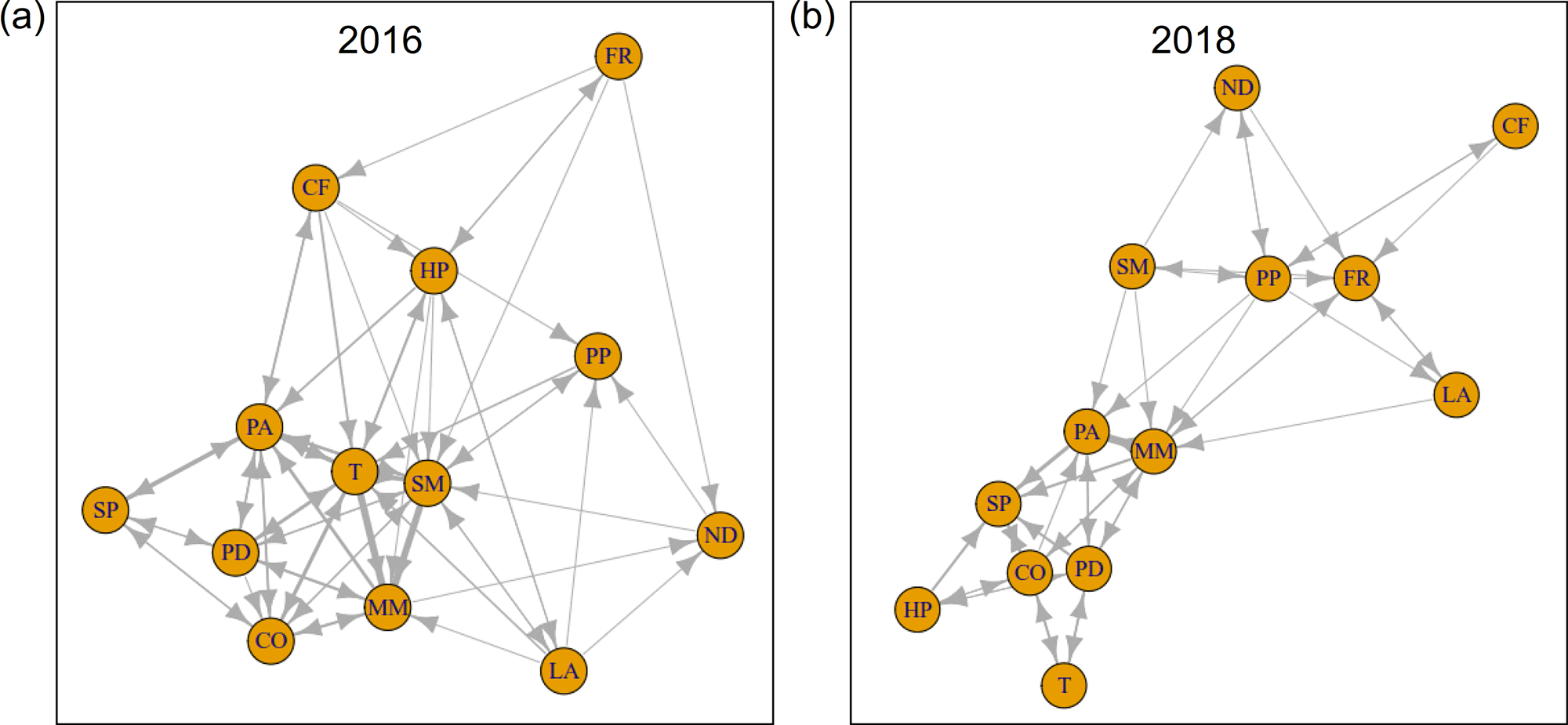
图9.2016年(a)和2018年(b)维管植物群落信息网络。边的宽度表示两个分量之间信息传递的强度。物种缩写在表1中展开。
表1
表1.1990 - 2018年第1地块记录的维管植物种类。
| 物种 | 缩写 |
| Cerastium fontanum | CF |
| Cochlearia officinalis | 有限公司 |
| 羊茅属rubra | FR |
| Honckenya peploides | 惠普 |
| Leymus arenarius | 拉 |
| Tripleurospermum maritimassp。maritima(同义词洋甘菊maritima) | 毫米 |
| 植物 | 巴勒斯坦权力机构 |
| Poa pratensis | 页 |
| Puccinellia distans | PD |
| Sagina procumbens | SP |
| Stellaria媒体 | SM |
| 蒲公英 | T |